

Accelerated disease progression and robust innate host response in aged SIVmac239-infected Chinese rhesus macaques is associated with enhanced immunosenescence
- PMID: 28232735
- PMCID: PMC5428349
- DOI: 10.1038/s41598-017-00084-0
Accelerated disease progression and robust innate host response in aged SIVmac239-infected Chinese rhesus macaques is associated with enhanced immunosenescence
Abstract
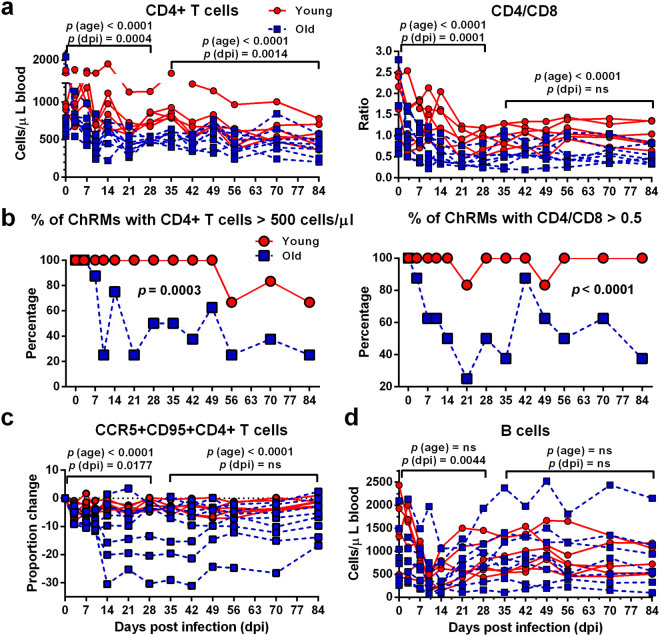
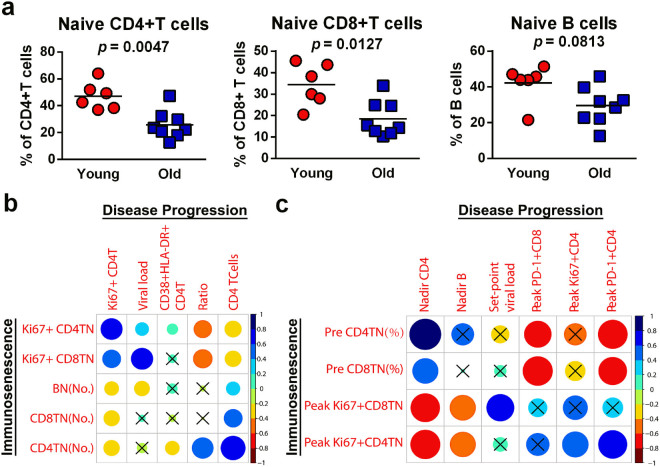
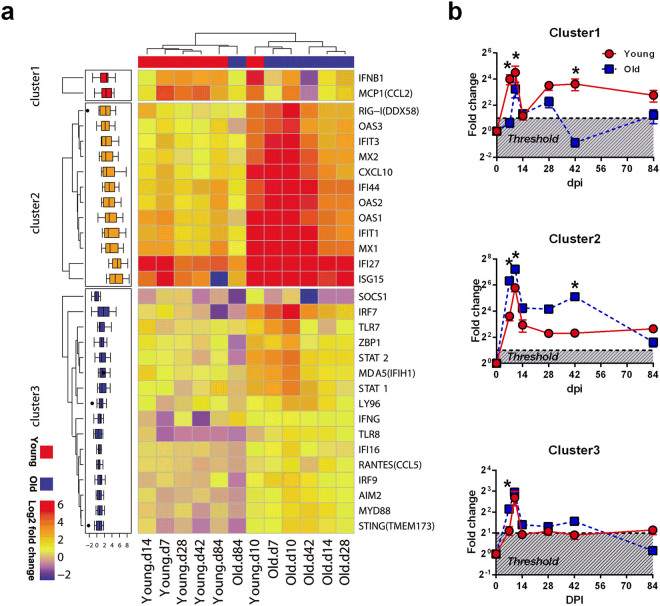
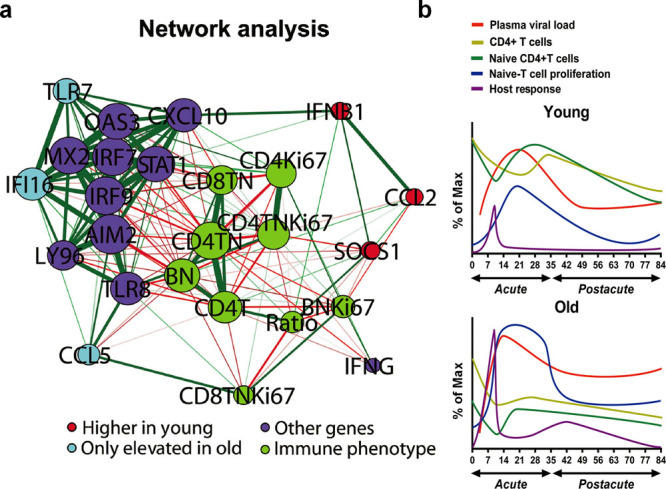
The elderly population infected with HIV-1 is often characterized by the rapid AIDS progression and poor treatment outcome, possibly because of immunosenescence resulting from both HIV infection and aging. However, this hypothesis remains to be fully tested. Here, we studied 6 young and 12 old Chinese rhesus macaques (ChRM) over the course of three months after simian immunodeficiency virus (SIV) SIVmac239 infection. Old ChRM showed a higher risk of accelerated AIDS development than did young macaques, owing to rapidly elevated plasma viral loads and decreased levels of CD4+ T cells. The low frequency of naïve CD4+ T cells before infection was strongly predictive of an increased disease progression, whereas the severe depletion of CD4+ T cells and the rapid proliferation of naïve lymphocytes accelerated the exhaustion of naïve lymphocytes in old ChRM. Moreover, in old ChRM, a robust innate host response with defective regulation was associated with a compensation for naïve T cell depletion and a high level of immune activation. Therefore, we suggest that immunosenescence plays an important role in the accelerated AIDS progression in elderly individuals and that SIV-infected old ChRM may be a favorable model for studying AIDS pathogenesis and researching therapies for elderly AIDS patients.
Figures



Similar articles
-
Critical Role for Monocytes/Macrophages in Rapid Progression to AIDS in Pediatric Simian Immunodeficiency Virus-Infected Rhesus Macaques.J Virol. 2017 Aug 10;91(17):e00379-17. doi: 10.1128/JVI.00379-17. Print 2017 Sep 1. J Virol. 2017. PMID: 28566378 Free PMC article.
-
Mycobacterium bovis bacille Calmette-Guérin enhances pathogenicity of simian immunodeficiency virus infection and accelerates progression to AIDS in macaques: a role of persistent T cell activation in AIDS pathogenesis.J Immunol. 1999 Feb 15;162(4):2204-16. J Immunol. 1999. PMID: 9973496
-
Loss of naïve cells accompanies memory CD4+ T-cell depletion during long-term progression to AIDS in Simian immunodeficiency virus-infected macaques.J Virol. 2007 Jan;81(2):893-902. doi: 10.1128/JVI.01635-06. Epub 2006 Nov 8. J Virol. 2007. PMID: 17093193 Free PMC article.
-
Monocytes in HIV and SIV Infection and Aging: Implications for Inflamm-Aging and Accelerated Aging.Viruses. 2022 Feb 17;14(2):409. doi: 10.3390/v14020409. Viruses. 2022. PMID: 35216002 Free PMC article. Review.
-
Mechanisms of immune aging in HIV.Clin Sci (Lond). 2022 Jan 14;136(1):61-80. doi: 10.1042/CS20210344. Clin Sci (Lond). 2022. PMID: 34985109 Review.
Cited by
-
Biomarkers of Activation and Inflammation to Track Disparity in Chronological and Physiological Age of People Living With HIV on Combination Antiretroviral Therapy.Front Immunol. 2020 Oct 9;11:583934. doi: 10.3389/fimmu.2020.583934. eCollection 2020. Front Immunol. 2020. PMID: 33162998 Free PMC article. Review.
-
Limited restoration of T cell subset distribution and immune function in older people living with HIV-1 receiving HAART.Immun Ageing. 2025 Jan 8;22(1):3. doi: 10.1186/s12979-024-00497-2. Immun Ageing. 2025. PMID: 39780181 Free PMC article.
-
Treatment outcomes amongst older people with HIV infection receiving antiretroviral therapy.AIDS. 2024 May 1;38(6):803-812. doi: 10.1097/QAD.0000000000003831. Epub 2024 Jan 12. AIDS. 2024. PMID: 38578958 Free PMC article.
-
Aging induces severe SIV infection accompanied by an increase in follicular CD8+ T cells with overactive STAT3 signaling.Cell Mol Immunol. 2022 Sep;19(9):1042-1053. doi: 10.1038/s41423-022-00899-6. Epub 2022 Jul 18. Cell Mol Immunol. 2022. PMID: 35851876 Free PMC article.
-
Type I interferon and HIV: Subtle balance between antiviral activity, immunopathogenesis and the microbiome.Cytokine Growth Factor Rev. 2018 Apr;40:19-31. doi: 10.1016/j.cytogfr.2018.03.003. Epub 2018 Mar 16. Cytokine Growth Factor Rev. 2018. PMID: 29576284 Free PMC article. Review.
References
-
- Kalayjian RC, et al. Age-related immune dysfunction in health and in human immunodeficiency virus (HIV) disease: association of age and HIV infection with naive CD8+ cell depletion, reduced expression of CD28 on CD8+ cells, and reduced thymic volumes. J Infect Dis. 2003;187:1924–1933. doi: 10.1086/375372. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
