

Genomic RNA folding mediates assembly of human parechovirus
- PMID: 28232749
- PMCID: PMC5431903
- DOI: 10.1038/s41467-016-0011-z
Genomic RNA folding mediates assembly of human parechovirus
Erratum in
-
Erratum: Genomic RNA folding mediates assembly of human parechovirus.Nat Commun. 2017 Jul 14;8(1):83. doi: 10.1038/s41467-017-00026-4. Nat Commun. 2017. PMID: 28710463 Free PMC article.
Abstract
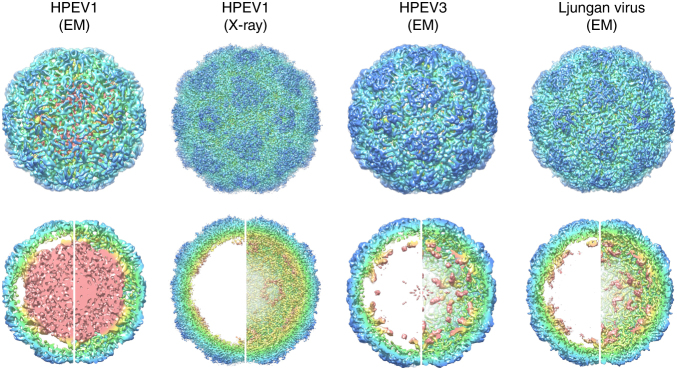
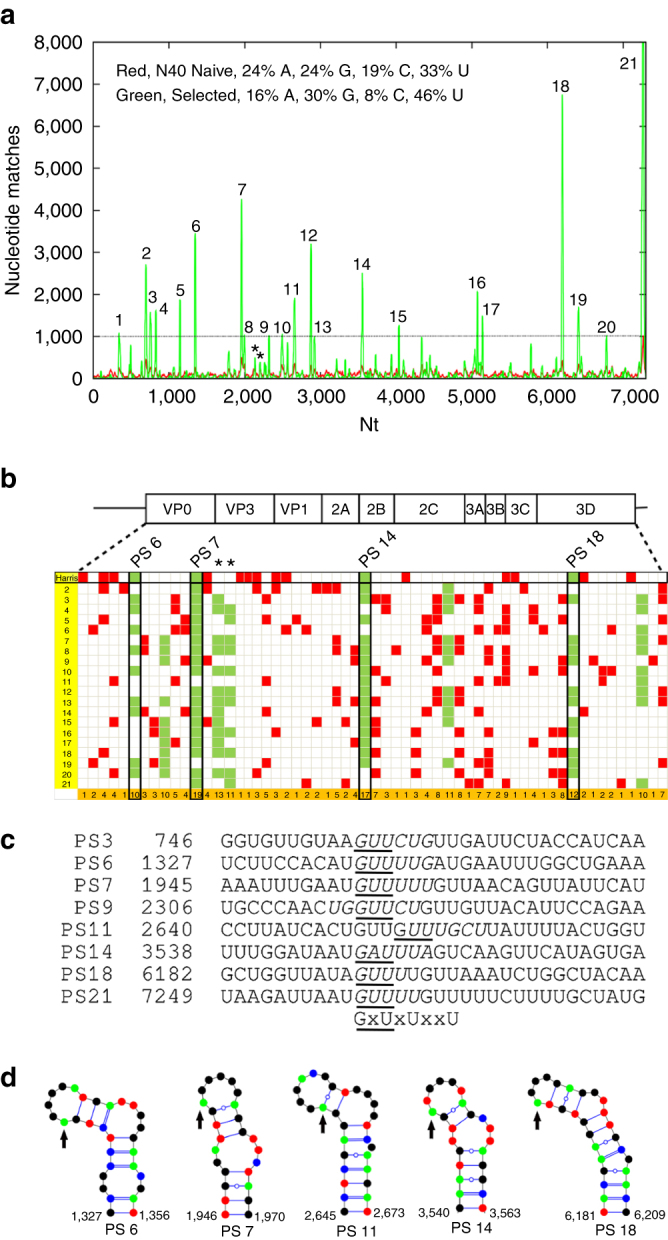
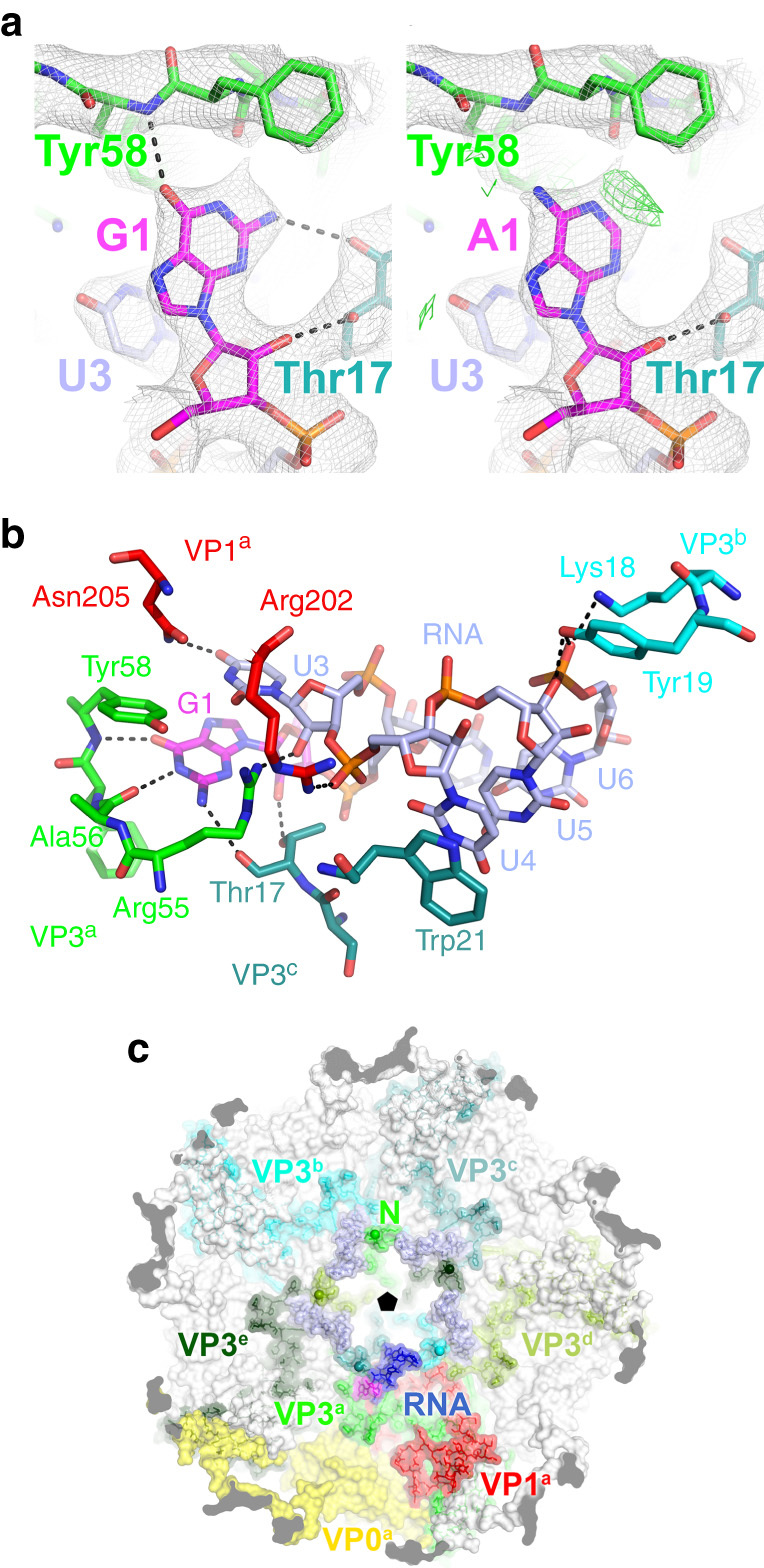
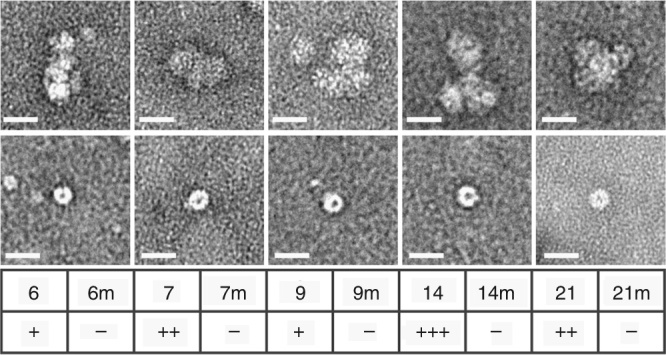
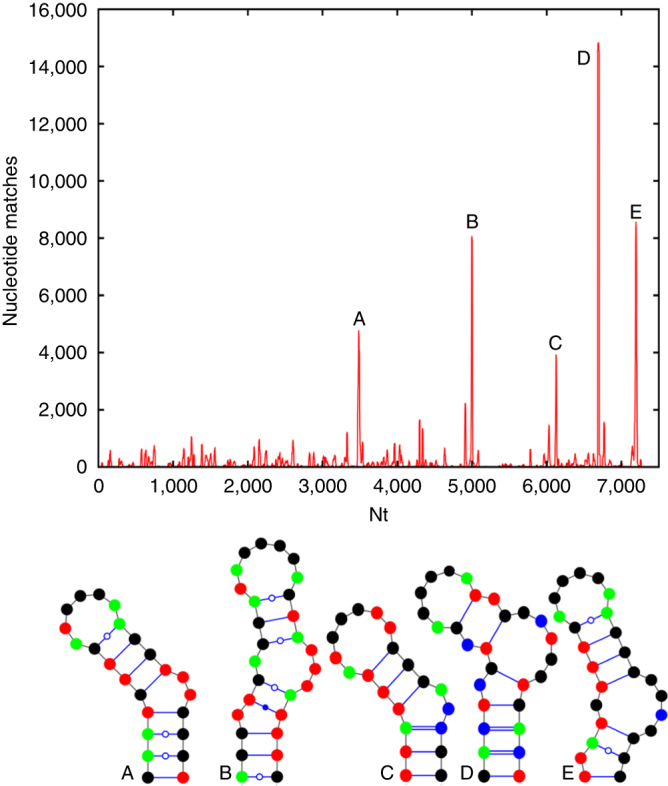
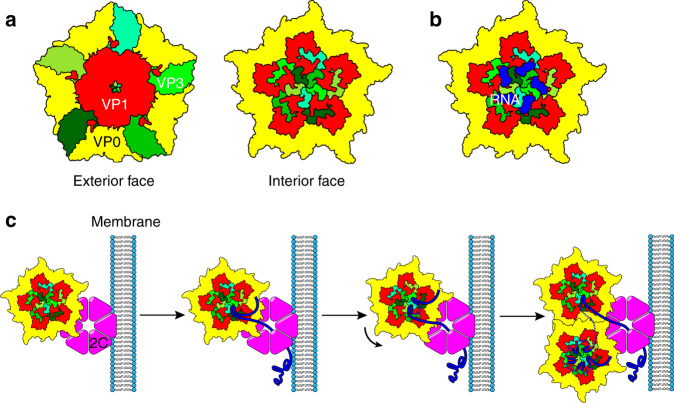
Assembly of the major viral pathogens of the Picornaviridae family is poorly understood. Human parechovirus 1 is an example of such viruses that contains 60 short regions of ordered RNA density making identical contacts with the protein shell. We show here via a combination of RNA-based systematic evolution of ligands by exponential enrichment, bioinformatics analysis and reverse genetics that these RNA segments are bound to the coat proteins in a sequence-specific manner. Disruption of either the RNA coat protein recognition motif or its contact amino acid residues is deleterious for viral assembly. The data are consistent with RNA packaging signals playing essential roles in virion assembly. Their binding sites on the coat proteins are evolutionarily conserved across the Parechovirus genus, suggesting that they represent potential broad-spectrum anti-viral targets.The mechanism underlying packaging of genomic RNA into viral particles is not well understood for human parechoviruses. Here the authors identify short RNA motifs in the parechovirus genome that bind capsid proteins, providing approximately 60 specific interactions for virion assembly.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
