

Pharmacological Characterization of Human Histamine Receptors and Histamine Receptor Mutants in the Sf9 Cell Expression System
- PMID: 28233175
- PMCID: PMC7120522
- DOI: 10.1007/164_2016_124
Pharmacological Characterization of Human Histamine Receptors and Histamine Receptor Mutants in the Sf9 Cell Expression System
Abstract
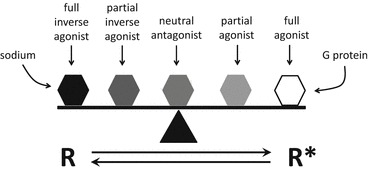
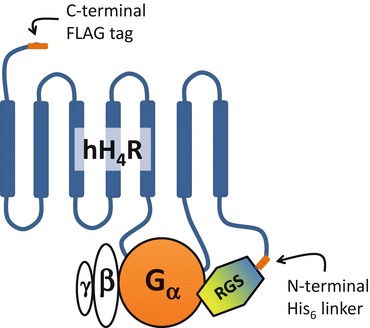
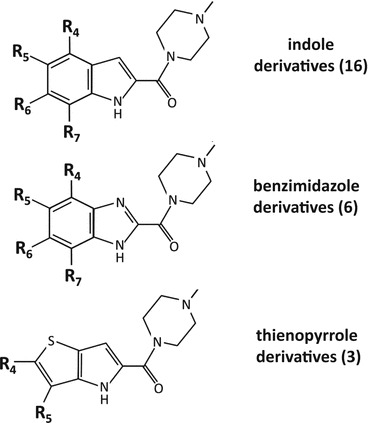
A large problem of histamine receptor research is data heterogeneity. Various experimental approaches, the complex signaling pathways of mammalian cells, and the use of different species orthologues render it difficult to compare and interpret the published results. Thus, the four human histamine receptor subtypes were analyzed side-by-side in the Sf9 insect cell expression system, using radioligand binding assays as well as functional readouts proximal to the receptor activation event (steady-state GTPase assays and [35S]GTPγS assays). The human H1R was co-expressed with the regulators of G protein signaling RGS4 or GAIP, which unmasked a productive interaction between hH1R and insect cell Gαq. By contrast, functional expression of the hH2R required the generation of an hH2R-Gsα fusion protein to ensure close proximity of G protein and receptor. Fusion of hH2R to the long (GsαL) or short (GsαS) splice variant of Gαs resulted in comparable constitutive hH2R activity, although both G protein variants show different GDP affinities. Medicinal chemistry studies revealed profound species differences between hH1R/hH2R and their guinea pig orthologues gpH1R/gpH2R. The causes for these differences were analyzed by molecular modeling in combination with mutational studies. Co-expression of the hH3R with Gαi1, Gαi2, Gαi3, and Gαi/o in Sf9 cells revealed high constitutive activity and comparable interaction efficiency with all G protein isoforms. A comparison of various cations (Li+, Na+, K+) and anions (Cl-, Br-, I-) revealed that anions with large radii most efficiently stabilize the inactive hH3R state. Potential sodium binding sites in the hH3R protein were analyzed by expressing specific hH3R mutants in Sf9 cells. In contrast to the hH3R, the hH4R preferentially couples to co-expressed Gαi2 in Sf9 cells. Its high constitutive activity is resistant to NaCl or GTPγS. The hH4R shows structural instability and adopts a G protein-independent high-affinity state. A detailed characterization of affinity and activity of a series of hH4R antagonists/inverse agonists allowed first conclusions about structure/activity relationships for inverse agonists at hH4R. In summary, the Sf9 cell system permitted a successful side-by-side comparison of all four human histamine receptor subtypes. This chapter summarizes the results of pharmacological as well as medicinal chemistry/molecular modeling approaches and demonstrates that these data are not only important for a deeper understanding of HxR pharmacology, but also have significant implications for the molecular pharmacology of GPCRs in general.
Keywords: GPCRs; Histamine receptors; Radioligand binding; Sf9 insect cells; Steady-state GTPase assay; [35S]GTPγS binding.
Figures



Similar articles
-
The human histamine H2-receptor couples more efficiently to Sf9 insect cell Gs-proteins than to insect cell Gq-proteins: limitations of Sf9 cells for the analysis of receptor/Gq-protein coupling.J Neurochem. 2002 Feb;80(4):678-96. doi: 10.1046/j.0022-3042.2001.00746.x. J Neurochem. 2002. PMID: 11841575
-
Distinct interaction of human and guinea pig histamine H2-receptor with guanidine-type agonists.Mol Pharmacol. 2001 Dec;60(6):1210-25. doi: 10.1124/mol.60.6.1210. Mol Pharmacol. 2001. PMID: 11723228
-
N1-(3-cyclohexylbutanoyl)-N2-[3-(1H-imidazol-4-yl)propyl]guanidine (UR-AK57), a potent partial agonist for the human histamine H1- and H2-receptors.J Pharmacol Exp Ther. 2006 Jun;317(3):1262-8. doi: 10.1124/jpet.106.102897. Epub 2006 Mar 22. J Pharmacol Exp Ther. 2006. PMID: 16554355
-
Sf9 cells: a versatile model system to investigate the pharmacological properties of G protein-coupled receptors.Pharmacol Ther. 2010 Dec;128(3):387-418. doi: 10.1016/j.pharmthera.2010.07.005. Epub 2010 Aug 10. Pharmacol Ther. 2010. PMID: 20705094 Review.
-
Molecular pharmacology of the four histamine receptors.Adv Exp Med Biol. 2010;709:11-9. doi: 10.1007/978-1-4419-8056-4_2. Adv Exp Med Biol. 2010. PMID: 21618883 Review.
Cited by
-
The Function of the Histamine H4 Receptor in Inflammatory and Inflammation-Associated Diseases of the Gut.Int J Mol Sci. 2021 Jun 6;22(11):6116. doi: 10.3390/ijms22116116. Int J Mol Sci. 2021. PMID: 34204101 Free PMC article. Review.
References
-
- Bahia DS, Sartania N, Ward RJ, Cavalli A, Jones TL, Druey KM, Milligan G. Concerted stimulation and deactivation of pertussis toxin-sensitive G proteins by chimeric G protein-coupled receptor-regulator of G protein signaling 4 fusion proteins: analysis of the contribution of palmitoylated cysteine residues to the GAP activity of RGS4. J Neurochem. 2003;85:1289–1298. doi: 10.1046/j.1471-4159.2003.01769.x. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
