

Increased 4R tau expression and behavioural changes in a novel MAPT-N296H genomic mouse model of tauopathy
- PMID: 28233851
- PMCID: PMC5324134
- DOI: 10.1038/srep43198
Increased 4R tau expression and behavioural changes in a novel MAPT-N296H genomic mouse model of tauopathy
Abstract
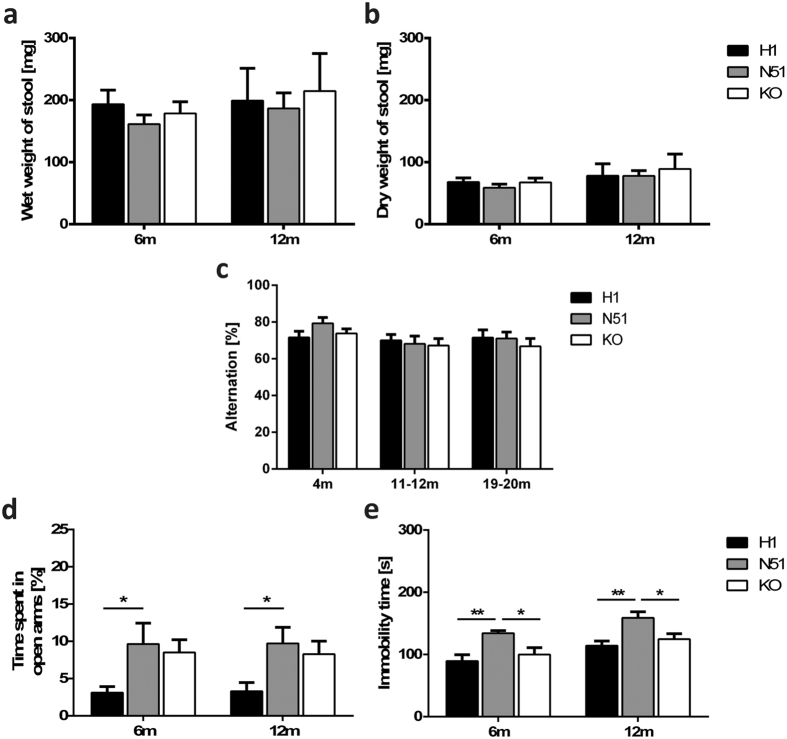
The microtubule-associated protein tau is implicated in various neurodegenerative diseases including Alzheimer's disease, progressive supranuclear palsy and corticobasal degeneration, which are characterized by intracellular accumulation of hyperphosphorylated tau. Mutations in the tau gene MAPT cause frontotemporal dementia with parkinsonism linked to chromosome 17 (FTDP-17). In the human central nervous system, six tau isoforms are expressed, and imbalances in tau isoform ratios are associated with pathology. To date, few animal models of tauopathy allow for the potential influence of these protein isoforms, relying instead on cDNA-based transgene expression. Using the P1-derived artificial chromosome (PAC) technology, we created mouse lines expressing all six tau isoforms from the human MAPT locus, harbouring either the wild-type sequence or the disease-associated N296H mutation on an endogenous Mapt-/- background. Animals expressing N296H mutant tau recapitulated early key features of tauopathic disease, including a tau isoform imbalance and tau hyperphosphorylation in the absence of somatodendritic tau inclusions. Furthermore, N296H animals displayed behavioural anomalies such as hyperactivity, increased time in the open arms of the elevated plus maze and increased immobility during the tail suspension test. The mouse models described provide an excellent model to study the function of wild-type or mutant tau in a highly physiological setting.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
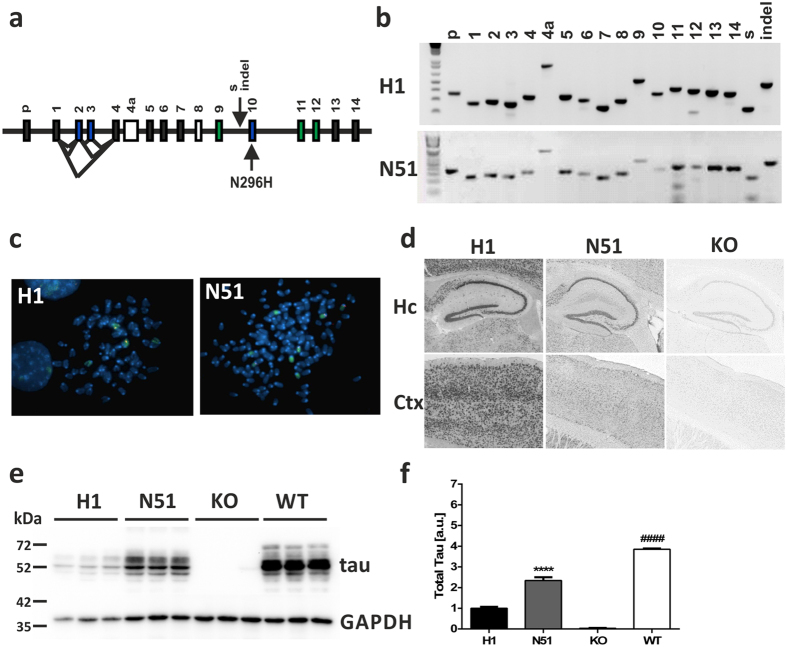
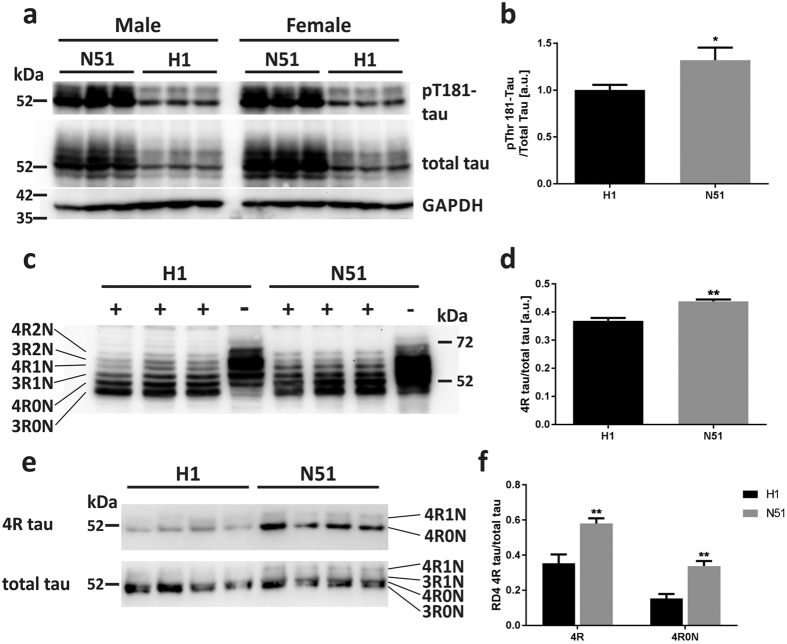
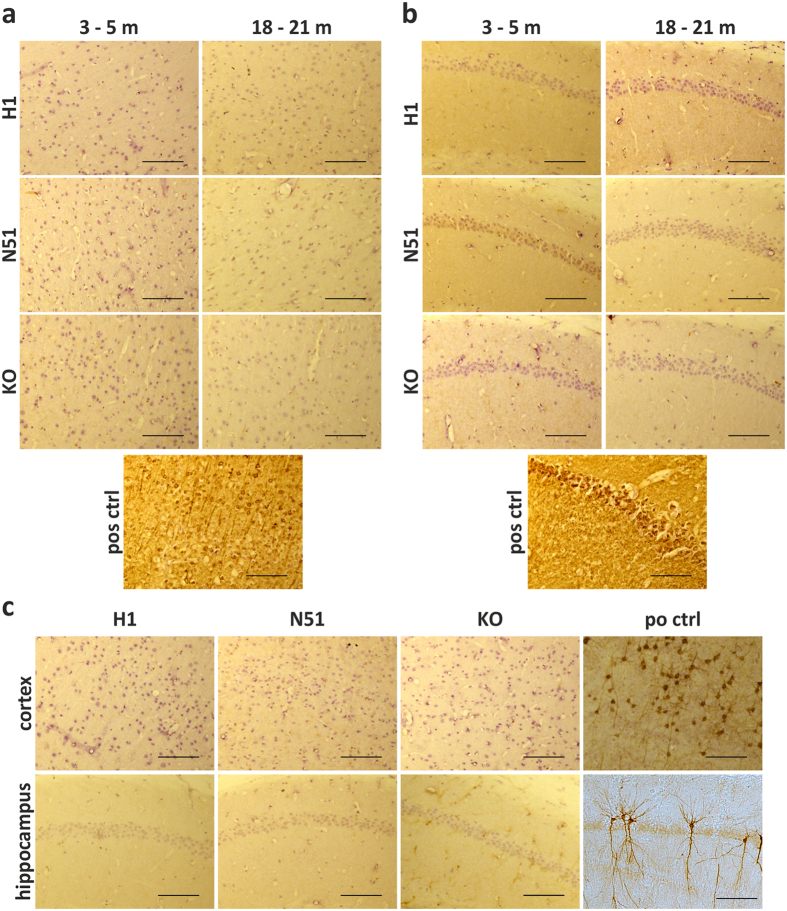
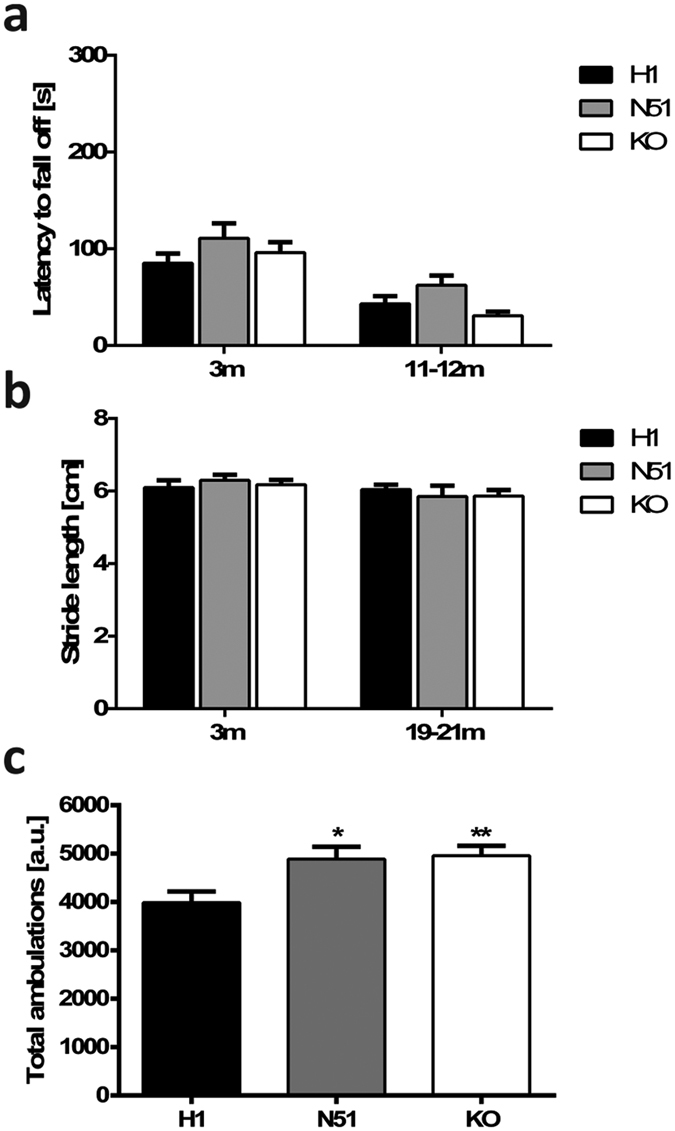
References
-
- Crawford F. et al.. No genetic association between polymorphisms in the Tau gene and Alzheimer’s disease in clinic or population based samples. Neurosci. Lett. 266, 193–196 (1999). - PubMed
-
- Myers A. J. et al.. The H1c haplotype at the MAPT locus is associated with Alzheimer’s disease. Hum. Mol. Genet. 14, 2399–2404 (2005). - PubMed
-
- Baker M. et al.. No association between TAU haplotype and Alzheimer’s disease in population or clinic based series or in familial disease. Neurosci. Lett. 285, 147–149 (2000). - PubMed
-
- Russ C. et al.. The microtubule associated protein Tau gene and Alzheimer’s disease–an association study and meta-analysis. Neurosci. Lett. 314, 92–96 (2001). - PubMed
-
- Abraham R. et al.. An association study of common variation at the MAPT locus with late-onset Alzheimer’s disease. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. Off. Publ. Int. Soc. Psychiatr. Genet. 150B, 1152–1155 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
