

Extra-epitopic hepatitis C virus polymorphisms confer resistance to broadly neutralizing antibodies by modulating binding to scavenger receptor B1
- PMID: 28235087
- PMCID: PMC5342271
- DOI: 10.1371/journal.ppat.1006235
Extra-epitopic hepatitis C virus polymorphisms confer resistance to broadly neutralizing antibodies by modulating binding to scavenger receptor B1
Abstract
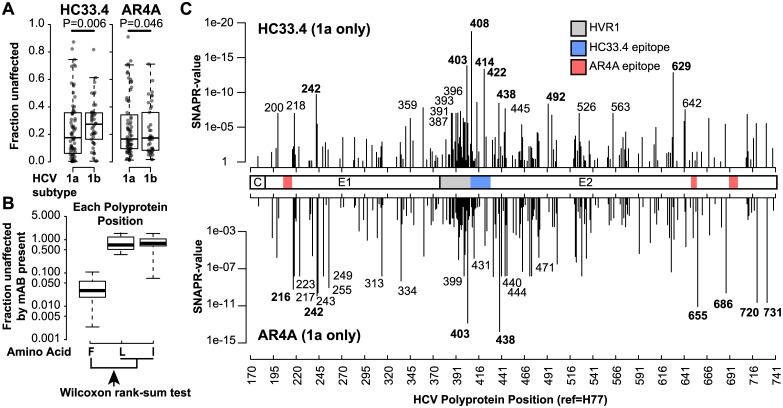
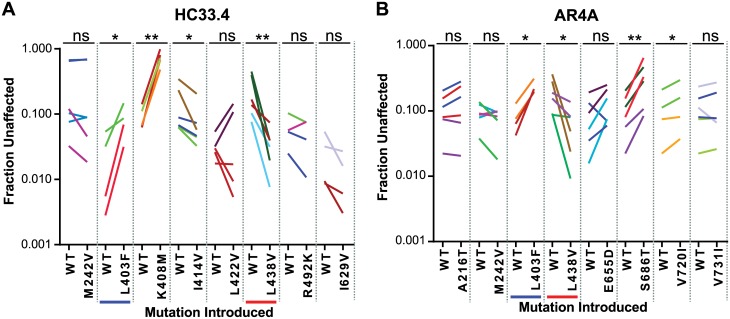
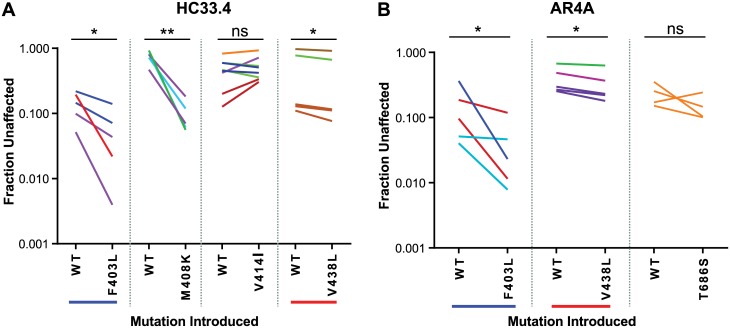
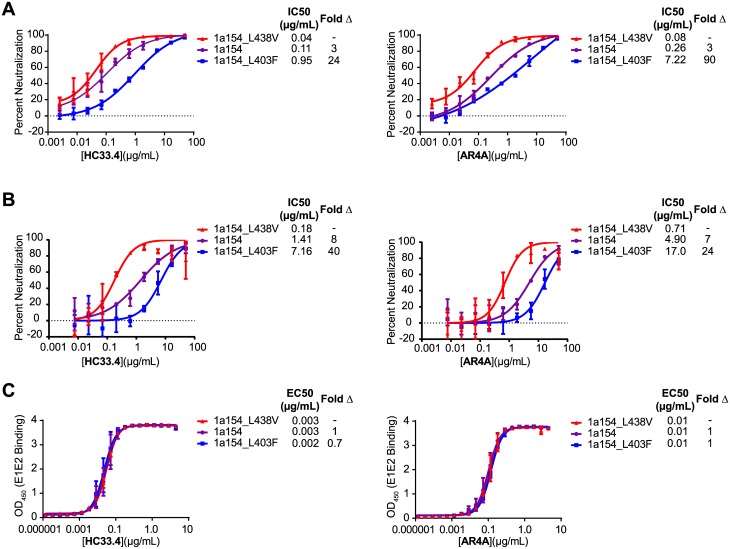
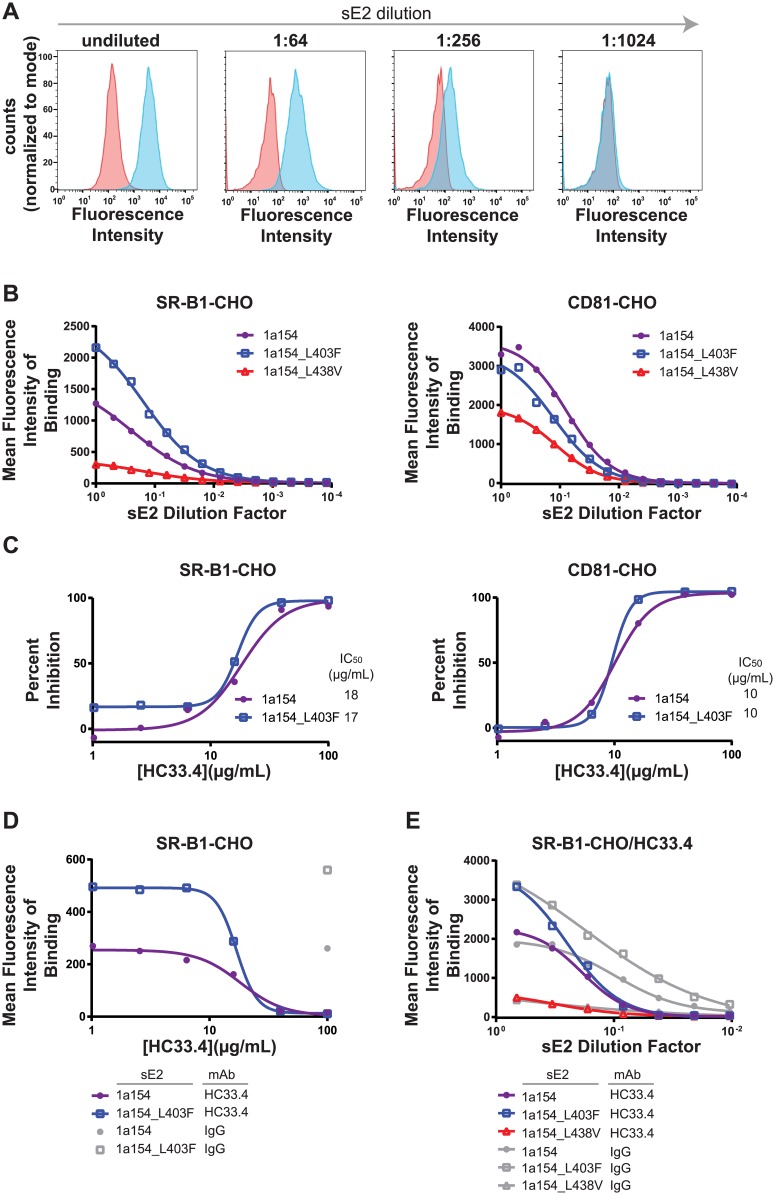
Broadly-neutralizing monoclonal antibodies (bNAbs) may guide vaccine development for highly variable viruses including hepatitis C virus (HCV), since they target conserved viral epitopes that could serve as vaccine antigens. However, HCV resistance to bNAbs could reduce the efficacy of a vaccine. HC33.4 and AR4A are two of the most potent anti-HCV human bNAbs characterized to date, binding to highly conserved epitopes near the amino- and carboxy-terminus of HCV envelope (E2) protein, respectively. Given their distinct epitopes, it was surprising that these bNAbs showed similar neutralization profiles across a panel of natural HCV isolates, suggesting that some viral polymorphisms may confer resistance to both bNAbs. To investigate this resistance, we developed a large, diverse panel of natural HCV envelope variants and a novel computational method to identify bNAb resistance polymorphisms in envelope proteins (E1 and E2). By measuring neutralization of a panel of HCV pseudoparticles by 10 μg/mL of each bNAb, we identified E1E2 variants with resistance to one or both bNAbs, despite 100% conservation of the AR4A binding epitope across the panel. We discovered polymorphisms outside of either binding epitope that modulate resistance to both bNAbs by altering E2 binding to the HCV co-receptor, scavenger receptor B1 (SR-B1). This study is focused on a mode of neutralization escape not addressed by conventional analysis of epitope conservation, highlighting the contribution of extra-epitopic polymorphisms to bNAb resistance and presenting a novel mechanism by which HCV might persist even in the face of an antibody response targeting multiple conserved epitopes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
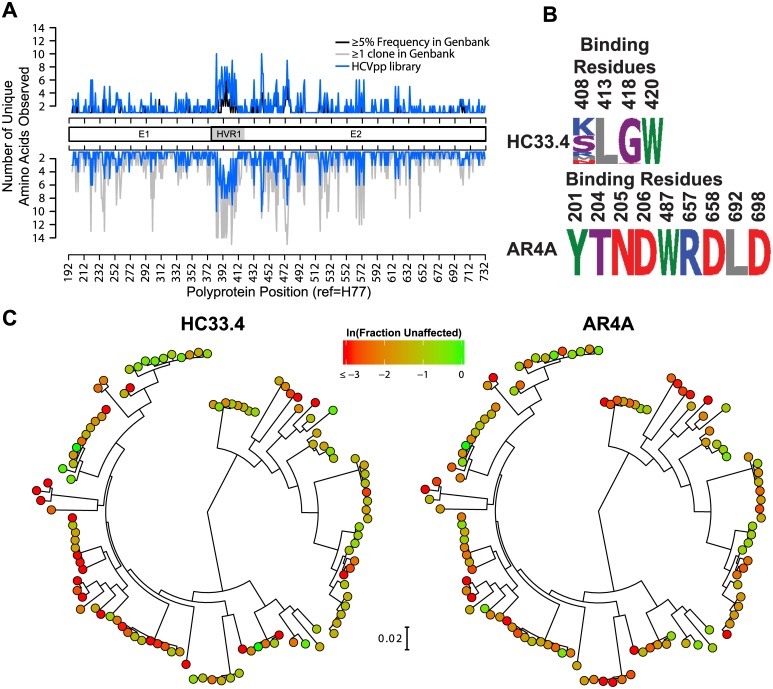
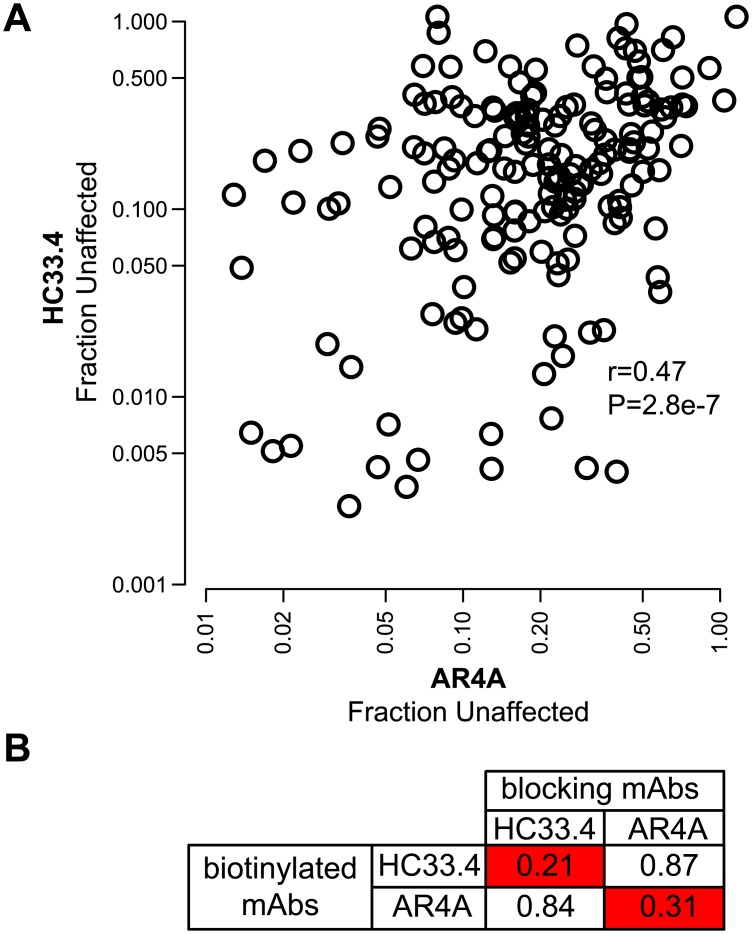
Similar articles
-
A Recombinant Hepatitis C Virus Genotype 1a E1/E2 Envelope Glycoprotein Vaccine Elicits Antibodies That Differentially Neutralize Closely Related 2a Strains through Interactions of the N-Terminal Hypervariable Region 1 of E2 with Scavenger Receptor B1.J Virol. 2019 Oct 29;93(22):e00810-19. doi: 10.1128/JVI.00810-19. Print 2019 Nov 15. J Virol. 2019. PMID: 31462563 Free PMC article.
-
Escape of Hepatitis C Virus from Epitope I Neutralization Increases Sensitivity of Other Neutralization Epitopes.J Virol. 2018 Apr 13;92(9):e02066-17. doi: 10.1128/JVI.02066-17. Print 2018 May 1. J Virol. 2018. PMID: 29467319 Free PMC article.
-
Structure-Based Design of Hepatitis C Virus E2 Glycoprotein Improves Serum Binding and Cross-Neutralization.J Virol. 2020 Oct 27;94(22):e00704-20. doi: 10.1128/JVI.00704-20. Print 2020 Oct 27. J Virol. 2020. PMID: 32878891 Free PMC article.
-
Mechanisms of HCV resistance to broadly neutralizing antibodies.Curr Opin Virol. 2021 Oct;50:23-29. doi: 10.1016/j.coviro.2021.07.003. Epub 2021 Jul 28. Curr Opin Virol. 2021. PMID: 34329953 Free PMC article. Review.
-
Defining Breadth of Hepatitis C Virus Neutralization.Front Immunol. 2018 Aug 2;9:1703. doi: 10.3389/fimmu.2018.01703. eCollection 2018. Front Immunol. 2018. PMID: 30116237 Free PMC article. Review.
Cited by
-
In vitro adaptation and characterization of attenuated hypervariable region 1 swap chimeras of hepatitis C virus.PLoS Pathog. 2021 Jul 19;17(7):e1009720. doi: 10.1371/journal.ppat.1009720. eCollection 2021 Jul. PLoS Pathog. 2021. PMID: 34280245 Free PMC article.
-
Hypervariable region 1 and N-linked glycans of hepatitis C regulate virion neutralization by modulating envelope conformations.Proc Natl Acad Sci U S A. 2019 May 14;116(20):10039-10047. doi: 10.1073/pnas.1822002116. Epub 2019 Apr 30. Proc Natl Acad Sci U S A. 2019. PMID: 31040211 Free PMC article.
-
Structure-Based and Rational Design of a Hepatitis C Virus Vaccine.Viruses. 2021 May 5;13(5):837. doi: 10.3390/v13050837. Viruses. 2021. PMID: 34063143 Free PMC article. Review.
-
Hypervariable Region 1 in Envelope Protein 2 of Hepatitis C Virus: A Linchpin in Neutralizing Antibody Evasion and Viral Entry.Front Immunol. 2018 Sep 27;9:2146. doi: 10.3389/fimmu.2018.02146. eCollection 2018. Front Immunol. 2018. PMID: 30319614 Free PMC article. Review.
-
Can Broadly Neutralizing Monoclonal Antibodies Lead to a Hepatitis C Virus Vaccine?Trends Microbiol. 2018 Oct;26(10):854-864. doi: 10.1016/j.tim.2018.04.002. Epub 2018 Apr 24. Trends Microbiol. 2018. PMID: 29703495 Free PMC article. Review.
References
-
- Thomas DL, Astemborski J, Rai RM, et al. The natural history of hepatitis c virus infection: Host, viral, and environmental factors. JAMA. 2000;284: 450–456. - PubMed
-
- Zibbell JE, Iqbal K, Patel RC, Suryaprasad A, Sanders KJ, Moore-Moravian L, et al. Increases in hepatitis C virus infection related to injection drug use among persons aged ≤30 years—kentucky, tennessee, virginia, and west virginia, 2006–2012. MMWR Morb Mortal Wkly Rep. 2015;64: 453–458. - PMC - PubMed
-
- Five–Year Risk of Late Relapse or Reinfection With Hepatitis C After Sustained Virological Response: Meta-analysis of 49 Studies in 8534 Patients [Internet]. [cited 7 Jun 2015]. http://www.croiconference.org/sessions/five%E2%80%93year-risk-late-relap...
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
