

A systems genetics approach identifies Trp53inp2 as a link between cardiomyocyte glucose utilization and hypertrophic response
- PMID: 28235788
- PMCID: PMC5407157
- DOI: 10.1152/ajpheart.00068.2016
A systems genetics approach identifies Trp53inp2 as a link between cardiomyocyte glucose utilization and hypertrophic response
Abstract
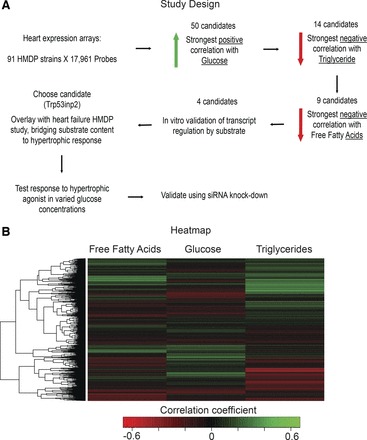
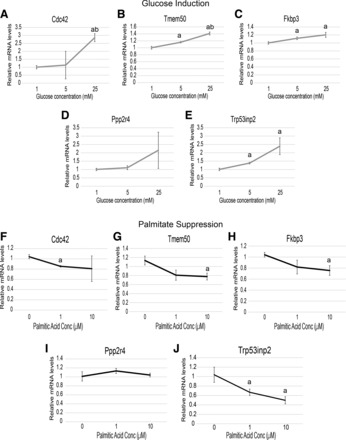
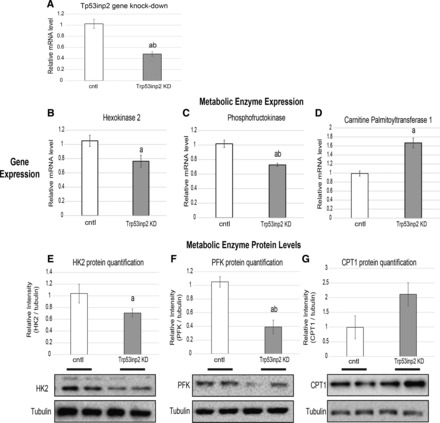
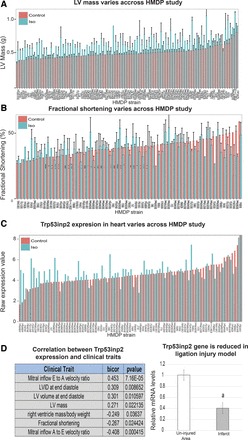
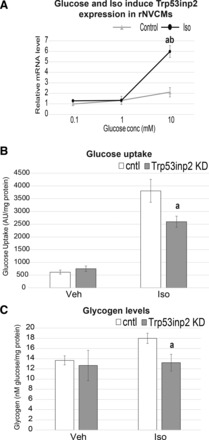
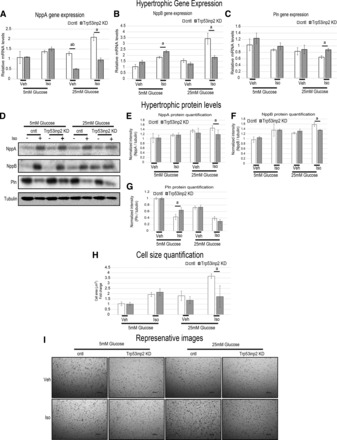
Cardiac failure has been widely associated with an increase in glucose utilization. The aim of our study was to identify factors that mechanistically bridge this link between hyperglycemia and heart failure. Here, we screened the Hybrid Mouse Diversity Panel (HMDP) for substrate-specific cardiomyocyte candidates based on heart transcriptional profile and circulating nutrients. Next, we utilized an in vitro model of rat cardiomyocytes to demonstrate that the gene expression changes were in direct response to substrate abundance. After overlaying candidates of interest with a separate HMDP study evaluating isoproterenol-induced heart failure, we chose to focus on the gene Trp53inp2 as a cardiomyocyte glucose utilization-specific factor. Trp53inp2 gene knockdown in rat cardiomyocytes reduced expression and protein abundance of key glycolytic enzymes. This resulted in reduction of both glucose uptake and glycogen content in cardiomyocytes stimulated with isoproterenol. Furthermore, this reduction effectively blunted the capacity of glucose and isoprotereonol to synergistically induce hypertrophic gene expression and cell size expansion. We conclude that Trp53inp2 serves as regulator of cardiomyocyte glycolytic activity and can consequently regulate hypertrophic response in the context of elevated glucose content.NEW & NOTEWORTHY Here, we apply a novel method for screening transcripts based on a substrate-specific expression pattern to identify Trp53inp2 as an induced cardiomyocyte glucose utilization factor. We further show that reducing expression of the gene could effectively blunt hypertrophic response in the context of elevated glucose content.
Keywords: Trp53inp2; glucose; hypertrophy; metabolic shift.
Copyright © 2017 the American Physiological Society.
Figures
Similar articles
-
Cardiomyocyte Proteome Remodeling due to Isoproterenol-Induced Cardiac Hypertrophy during the Compensated Phase.Proteomics Clin Appl. 2020 Jul;14(4):e2000017. doi: 10.1002/prca.202000017. Epub 2020 Jun 23. Proteomics Clin Appl. 2020. PMID: 32506788
-
Protocatechuic aldehyde protects against isoproterenol-induced cardiac hypertrophy via inhibition of the JAK2/STAT3 signaling pathway.Naunyn Schmiedebergs Arch Pharmacol. 2018 Dec;391(12):1373-1385. doi: 10.1007/s00210-018-1556-7. Epub 2018 Aug 21. Naunyn Schmiedebergs Arch Pharmacol. 2018. PMID: 30132020
-
The orphan receptor NOR1 participates in isoprenaline-induced cardiac hypertrophy by regulating PARP-1.Br J Pharmacol. 2015 Jun;172(11):2852-63. doi: 10.1111/bph.13091. Epub 2015 Mar 26. Br J Pharmacol. 2015. PMID: 25625556 Free PMC article.
-
Effects and mechanism of action of ligustrazine on isoprenaline-induced cardiomyocyte hypertrophy.Cell Biochem Biophys. 2014 Dec;70(3):1513-8. doi: 10.1007/s12013-014-0086-2. Cell Biochem Biophys. 2014. PMID: 25027096
-
MicroRNA-22 regulates cardiac hypertrophy and remodeling in response to stress.Circ Res. 2013 Apr 26;112(9):1234-43. doi: 10.1161/CIRCRESAHA.112.300682. Epub 2013 Mar 22. Circ Res. 2013. PMID: 23524588 Free PMC article.
Cited by
-
The potential of integrating human and mouse discovery platforms to advance our understanding of cardiometabolic diseases.Elife. 2023 Mar 31;12:e86139. doi: 10.7554/eLife.86139. Elife. 2023. PMID: 37000167 Free PMC article.
-
Metabolic status differentiates Trp53inp2 function in pressure-overload induced heart failure.Front Cardiovasc Med. 2023 Dec 18;10:1226586. doi: 10.3389/fcvm.2023.1226586. eCollection 2023. Front Cardiovasc Med. 2023. PMID: 38188257 Free PMC article.
-
Sex-specific genetic regulation of adipose mitochondria and metabolic syndrome by Ndufv2.Nat Metab. 2021 Nov;3(11):1552-1568. doi: 10.1038/s42255-021-00481-w. Epub 2021 Oct 25. Nat Metab. 2021. PMID: 34697471 Free PMC article.
-
Genetic architecture of heart mitochondrial proteome influencing cardiac hypertrophy.Elife. 2023 Jun 5;12:e82619. doi: 10.7554/eLife.82619. Elife. 2023. PMID: 37276142 Free PMC article.
-
Guidelines for measuring cardiac physiology in mice.Am J Physiol Heart Circ Physiol. 2018 Apr 1;314(4):H733-H752. doi: 10.1152/ajpheart.00339.2017. Epub 2018 Jan 5. Am J Physiol Heart Circ Physiol. 2018. PMID: 29351456 Free PMC article.
References
-
- Allard MF, Schönekess BO, Henning SL, English DR, Lopaschuk GD. Contribution of oxidative metabolism and glycolysis to ATP production in hypertrophied hearts. Am J Physiol Heart Circ Physiol 267: H742–H750, 1994. - PubMed
-
- Amorim PA, Nguyen TD, Shingu Y, Schwarzer M, Mohr FW, Schrepper A, Doenst T. Myocardial infarction in rats causes partial impairment in insulin response associated with reduced fatty acid oxidation and mitochondrial gene expression. J Thorac Cardiovasc Surg 140: 1160–1167, 2010. doi:10.1016/j.jtcvs.2010.08.003. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources