

Biochemical characterization and structure determination of a potent, selective antibody inhibitor of human MMP9
- PMID: 28235803
- PMCID: PMC5399127
- DOI: 10.1074/jbc.M116.760579
Biochemical characterization and structure determination of a potent, selective antibody inhibitor of human MMP9
Abstract
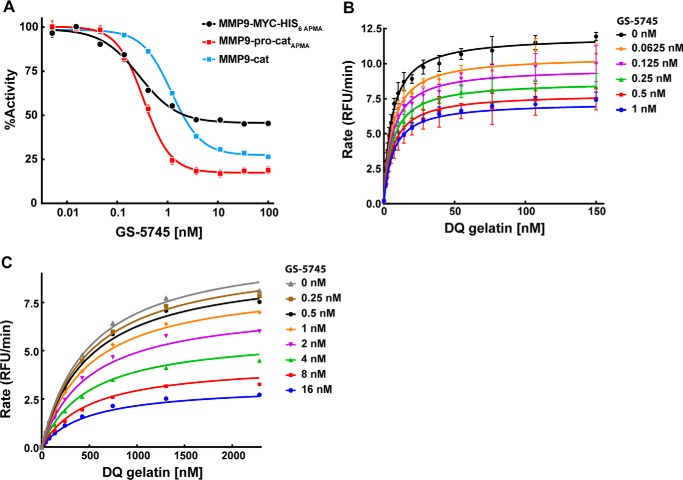
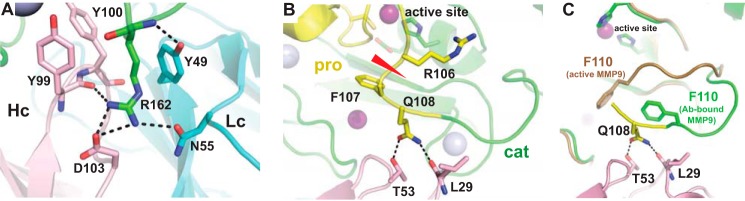
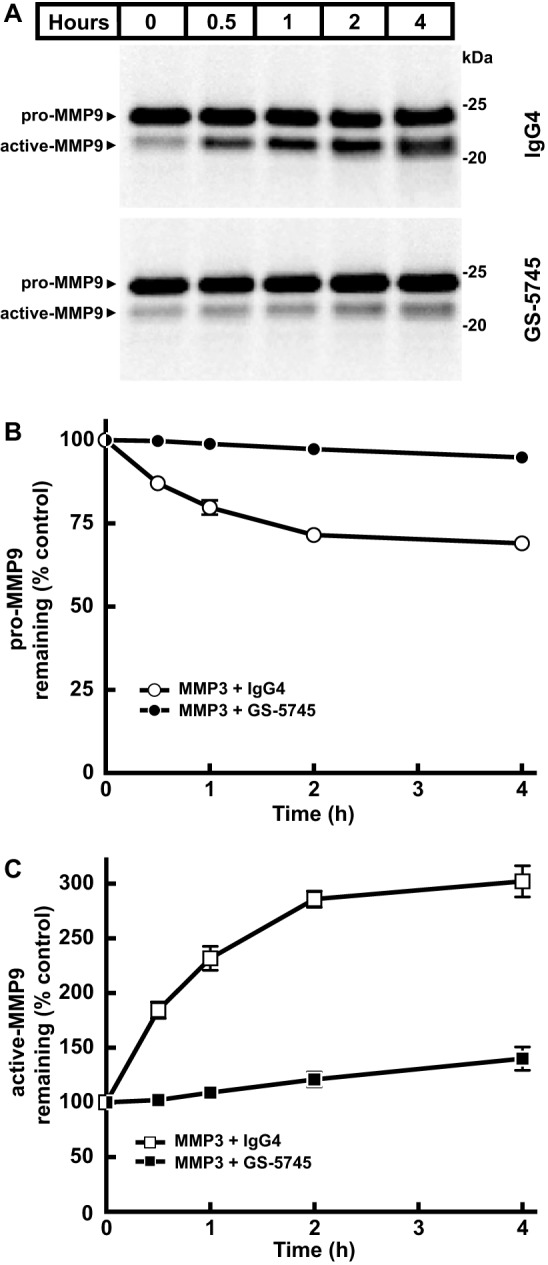
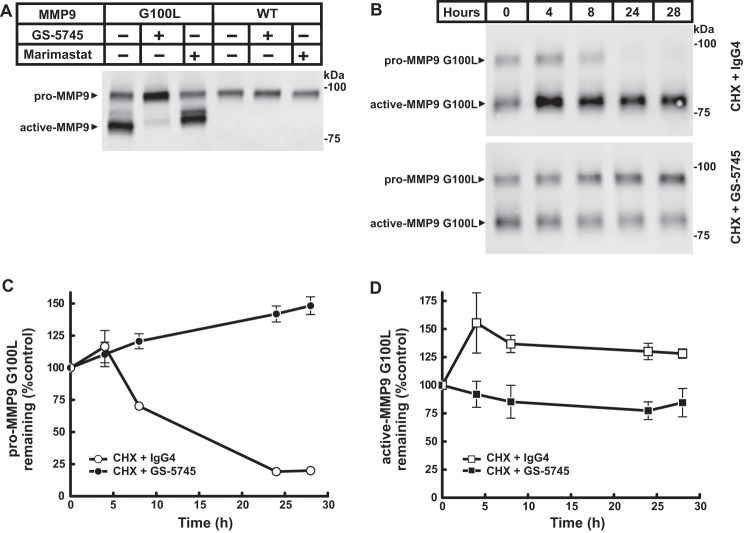
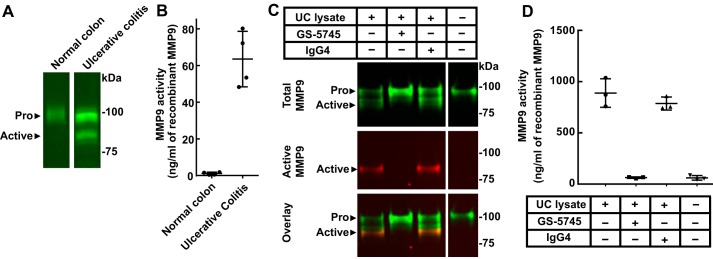
Matrix metalloproteinase 9 (MMP9) is a member of a large family of proteases that are secreted as inactive zymogens. It is a key regulator of the extracellular matrix, involved in the degradation of various extracellular matrix proteins. MMP9 plays a pathological role in a variety of inflammatory and oncology disorders and has long been considered an attractive therapeutic target. GS-5745, a potent, highly selective humanized monoclonal antibody inhibitor of MMP9, has shown promise in treating ulcerative colitis and gastric cancer. Here we describe the crystal structure of GS-5745·MMP9 complex and biochemical studies to elucidate the mechanism of inhibition of MMP9 by GS-5745. GS-5745 binds MMP9 distal to the active site, near the junction between the prodomain and catalytic domain, and inhibits MMP9 by two mechanisms. Binding to pro-MMP9 prevents MMP9 activation, whereas binding to active MMP9 allosterically inhibits activity.
Keywords: X-ray crystallography; allosteric regulation; antibody; enzyme inhibitor; matrix metalloproteinase (MMP).
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
All authors are employees of Gilead Sciences, Inc.
Figures

References
-
- Nagase H., and Woessner J. F. Jr. (1999) Matrix metalloproteinases. J. Biol. Chem. 274, 21491–21494 - PubMed
-
- Elkins P. A., Ho Y. S., Smith W. W., Janson C. A., D'Alessio K. J., McQueney M. S., Cummings M. D., and Romanic A. M. (2002) Structure of the C-terminally truncated human proMMP9, a gelatin-binding matrix metalloproteinase. Acta Crystallogr. D Biol. Crystallogr. 58, 1182–1192 - PubMed
-
- Vandooren J., Van den Steen P. E., and Opdenakker G. (2013) Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): the next decade. Crit. Rev. Biochem. Mol. Biol. 48, 222–272 - PubMed
-
- Ogata Y., Enghild J. J., and Nagase H. (1992) Matrix metalloproteinase 3 (stromelysin) activates the precursor for the human matrix metalloproteinase 9. J. Biol. Chem. 267, 3581–3584 - PubMed
-
- Shapiro S. D., Fliszar C. J., Broekelmann T. J., Mecham R. P., Senior R. M., and Welgus H. G. (1995) Activation of the 92-kDa gelatinase by stromelysin and 4-aminophenylmercuric acetate. Differential processing and stabilization of the carboxyl-terminal domain by tissue inhibitor of metalloproteinases (TIMP). J. Biol. Chem. 270, 6351–6356 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
