

A Novel Subfamily Esterase with a Homoserine Transacetylase-like Fold but No Transferase Activity
- PMID: 28235874
- PMCID: PMC5394324
- DOI: 10.1128/AEM.00131-17
A Novel Subfamily Esterase with a Homoserine Transacetylase-like Fold but No Transferase Activity
Abstract
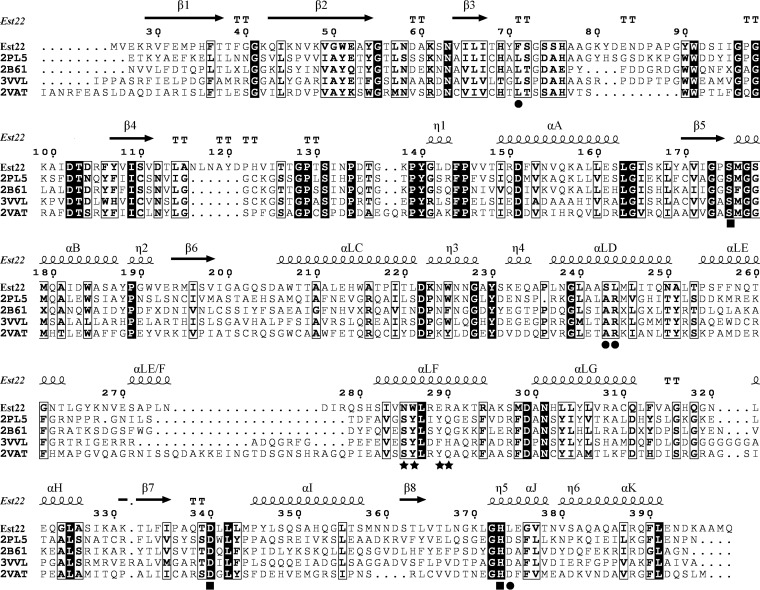
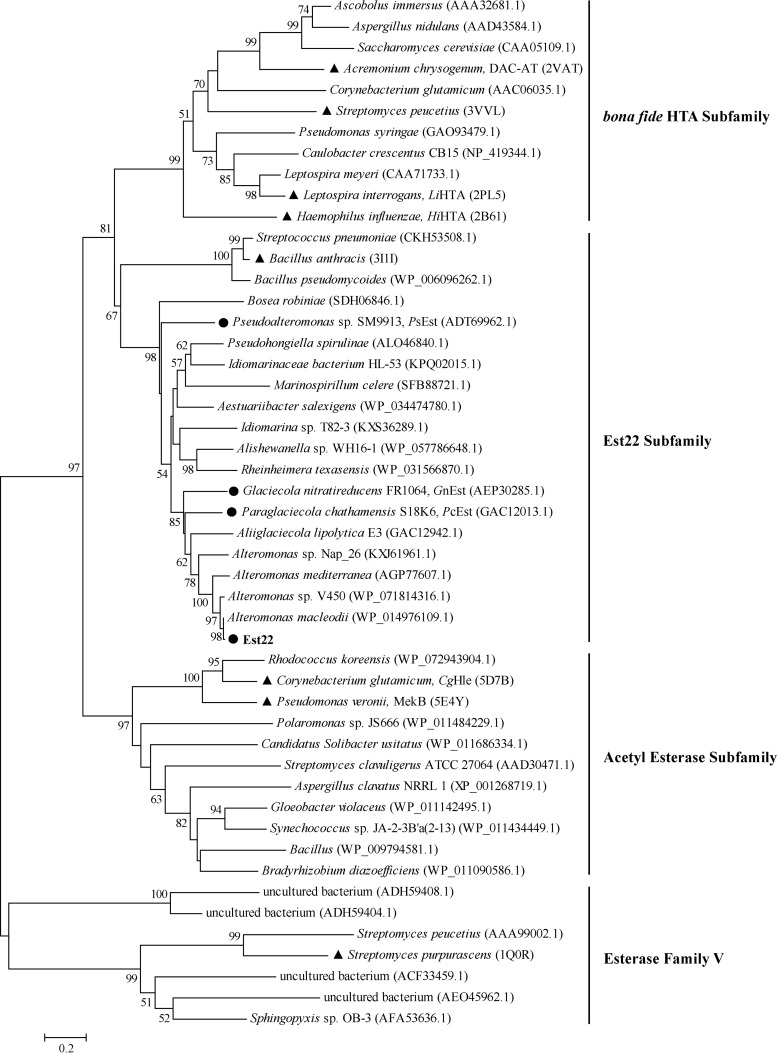
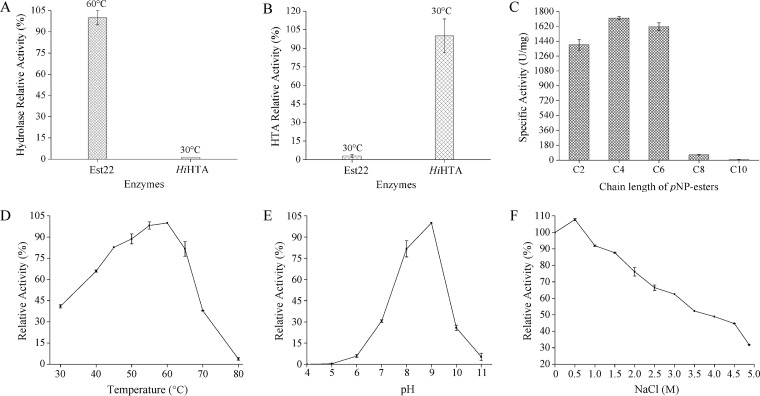
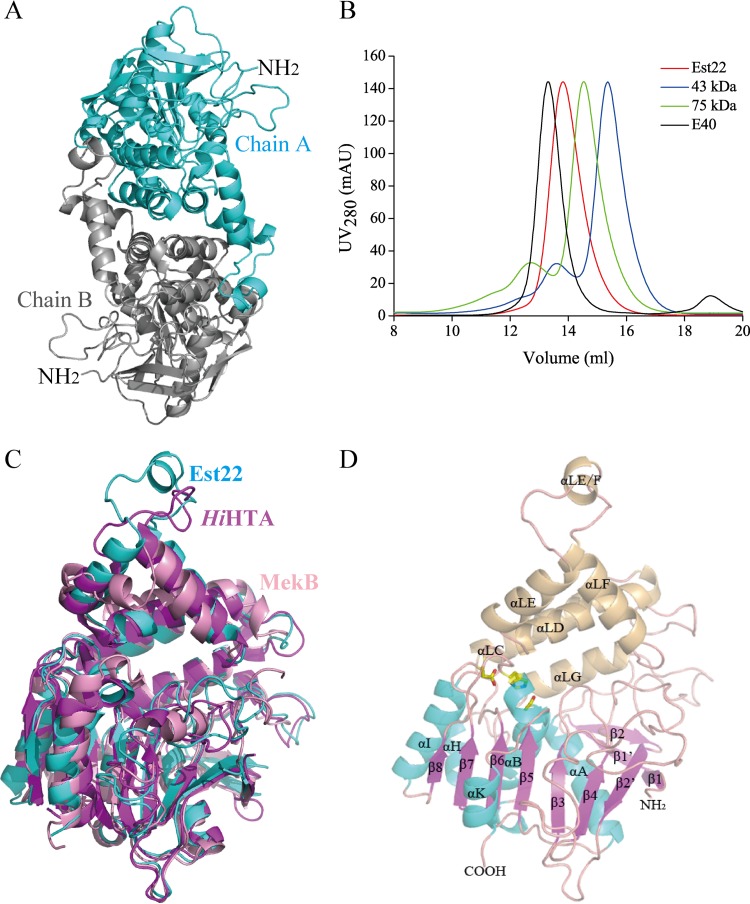
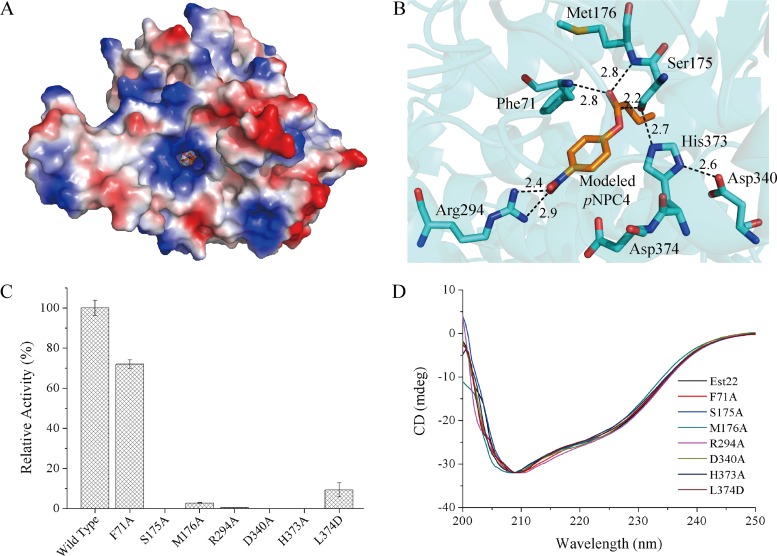
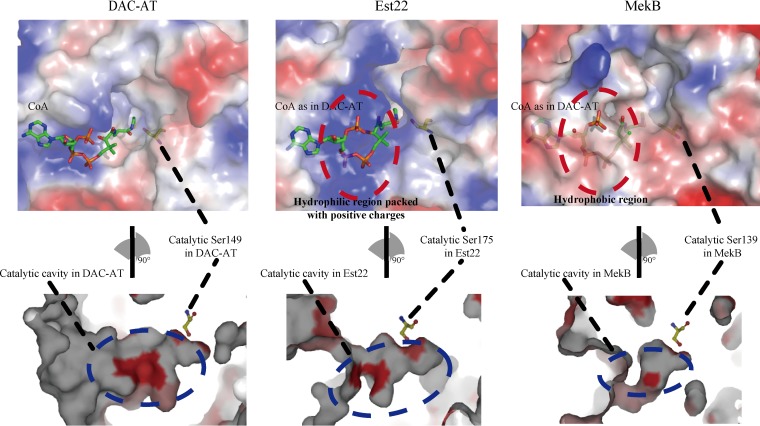
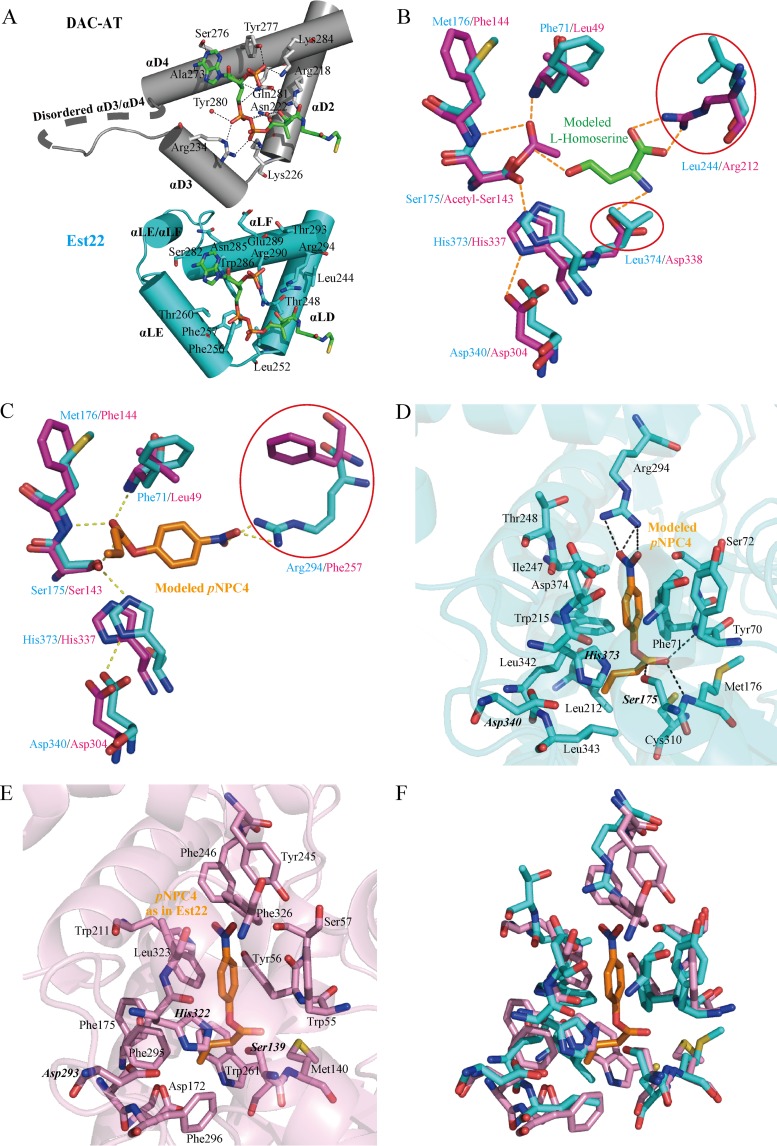
Microbial esterases play important roles in deep-sea organic carbon degradation and cycling. Although they have similar catalytic triads and oxyanion holes, esterases are hydrolases and homoserine transacetylases (HTAs) are transferases. Because two HTA homologs were identified as acetyl esterases, the HTA family was recently divided into the bona fide acetyltransferase subfamily and the acetyl esterase subfamily. Here, we identified and characterized a novel HTA-like esterase, Est22, from a deep-sea sedimentary metagenomic library. Est22 could efficiently hydrolyze esters with acyl lengths of up to six carbon atoms but had no transacetylase activity, which is different from HTAs and HTA-like acetyl esterases. Phylogenetic analysis also showed that Est22 and its homologs form a separate branch of the HTA family. We solved the structures of Est22 and its L374D mutant and modeled the structure of the L374D mutant with p-nitrophenyl butyrate. Based on structural, mutational, and biochemical analyses, Phe71 and Met176 in the oxyanion hole and Arg294 were revealed to be the key substrate-binding residues. A detailed structural comparison indicated that differences in their catalytic tunnels lead to the different substrate specificities of Est22 and the other two HTA subfamilies. Biochemical and sequence analyses suggested that Est22 homologs may have the same substrate recognition and catalysis mechanisms as Est22. Due to the significant differences in sequences, structures, and substrate specificities between Est22 (and its homologs) and the other two HTA subfamilies, we suggest that Est22 and its homologs represent a new subfamily in the HTA family.IMPORTANCE Microbial esterases play important roles in the turnover of organic carbon in the deep sea. Esterases and HTAs represent two groups of α/β hydrolases. Esterases catalyze the hydrolysis of simple esters and are widely used in the pharmaceutical and agrochemical industries, while HTAs catalyze the transfer of an acetyl group from acetyl-coenzyme A (CoA) to homoserine and are essential for microbial growth. Here, we report on a novel HTA-like esterase, Est22, from a deep-sea sediment. Because of the significant differences in sequences, structures, and substrate specificities of HTAs and HTA-like acetyl esterases, Est22 and its homologs represent a new subfamily in the HTA family. This study offers new knowledge regarding marine esterases.
Keywords: catalysis; crystal structure; new HTA subfamily; serine esterase; substrate recognition.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
A novel esterase subfamily with α/β-hydrolase fold suggested by structures of two bacterial enzymes homologous to L-homoserine O-acetyl transferases.FEBS Lett. 2016 Jan;590(1):174-84. doi: 10.1002/1873-3468.12031. Epub 2015 Dec 28. FEBS Lett. 2016. PMID: 26787467
-
Two novel deep-sea sediment metagenome-derived esterases: residue 199 is the determinant of substrate specificity and preference.Microb Cell Fact. 2018 Jan 30;17(1):16. doi: 10.1186/s12934-018-0864-4. Microb Cell Fact. 2018. PMID: 29382330 Free PMC article.
-
Functional characterisation of a metagenome derived family VIII esterase with a deacetylation activity on β-lactam antibiotics.Biochem Biophys Res Commun. 2013 Aug 2;437(3):342-8. doi: 10.1016/j.bbrc.2013.06.076. Epub 2013 Jul 1. Biochem Biophys Res Commun. 2013. PMID: 23827391
-
GDSL family of serine esterases/lipases.Prog Lipid Res. 2004 Nov;43(6):534-52. doi: 10.1016/j.plipres.2004.09.002. Prog Lipid Res. 2004. PMID: 15522763 Review.
-
Distinction between esterases and lipases: comparative biochemical properties of sequence-related carboxylesterases.Protein Pept Lett. 2009;16(10):1149-61. doi: 10.2174/092986609789071333. Protein Pept Lett. 2009. PMID: 19508178 Review.
Cited by
-
The FilZ Protein Contains a Single PilZ Domain and Facilitates the Swarming Motility of Pseudoalteromonas sp. SM9913.Microorganisms. 2023 Jun 13;11(6):1566. doi: 10.3390/microorganisms11061566. Microorganisms. 2023. PMID: 37375068 Free PMC article.
-
Excellent Degradation Performance of a Versatile Phthalic Acid Esters-Degrading Bacterium and Catalytic Mechanism of Monoalkyl Phthalate Hydrolase.Int J Mol Sci. 2018 Sep 18;19(9):2803. doi: 10.3390/ijms19092803. Int J Mol Sci. 2018. PMID: 30231475 Free PMC article.
-
Metagenomic Screening for Lipolytic Genes Reveals an Ecology-Clustered Distribution Pattern.Front Microbiol. 2022 Jun 10;13:851969. doi: 10.3389/fmicb.2022.851969. eCollection 2022. Front Microbiol. 2022. PMID: 35756004 Free PMC article.
-
A Novel and Efficient Phthalate Hydrolase from Acinetobacter sp. LUNF3: Molecular Cloning, Characterization and Catalytic Mechanism.Molecules. 2023 Sep 21;28(18):6738. doi: 10.3390/molecules28186738. Molecules. 2023. PMID: 37764514 Free PMC article.
-
A De Novo Designed Esterase with p-Nitrophenyl Acetate Hydrolysis Activity.Molecules. 2020 Oct 13;25(20):4658. doi: 10.3390/molecules25204658. Molecules. 2020. PMID: 33066055 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
