

Evaluation of a novel multi-immunogen vaccine strategy for targeting 4E10/10E8 neutralizing epitopes on HIV-1 gp41 membrane proximal external region
- PMID: 28237764
- PMCID: PMC5385849
- DOI: 10.1016/j.virol.2017.02.015
Evaluation of a novel multi-immunogen vaccine strategy for targeting 4E10/10E8 neutralizing epitopes on HIV-1 gp41 membrane proximal external region
Abstract
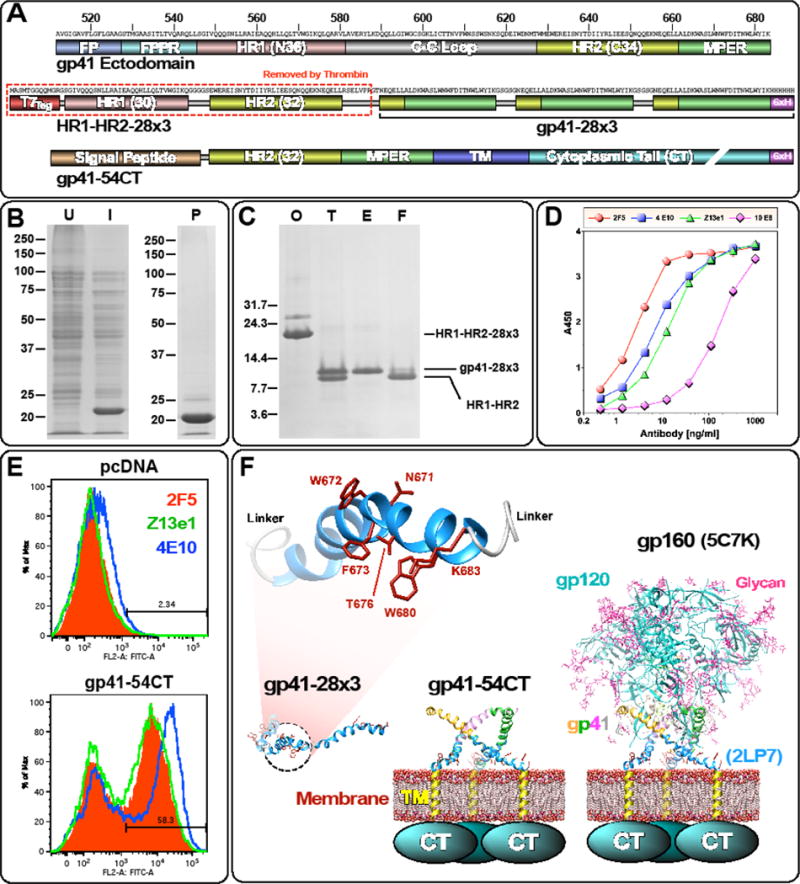
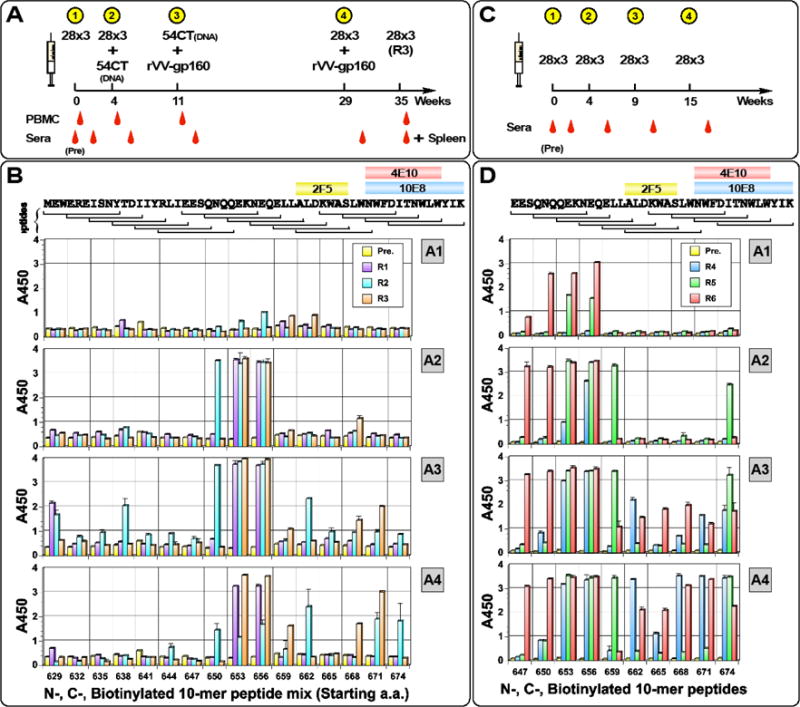
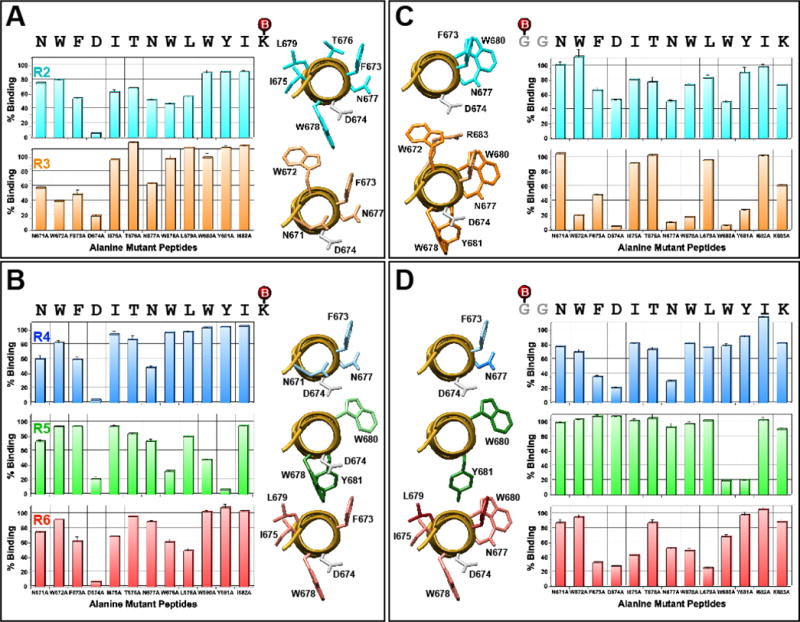
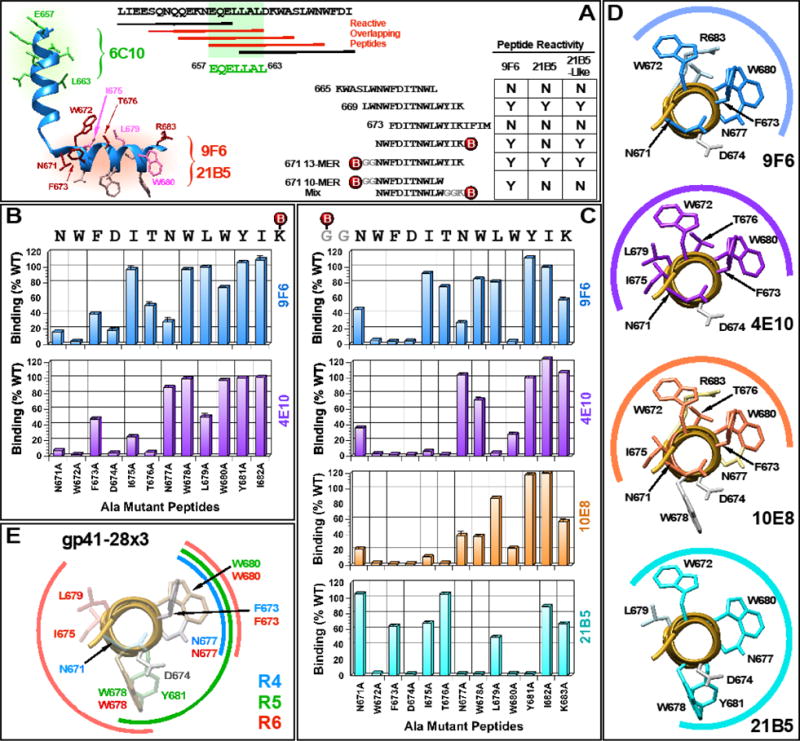
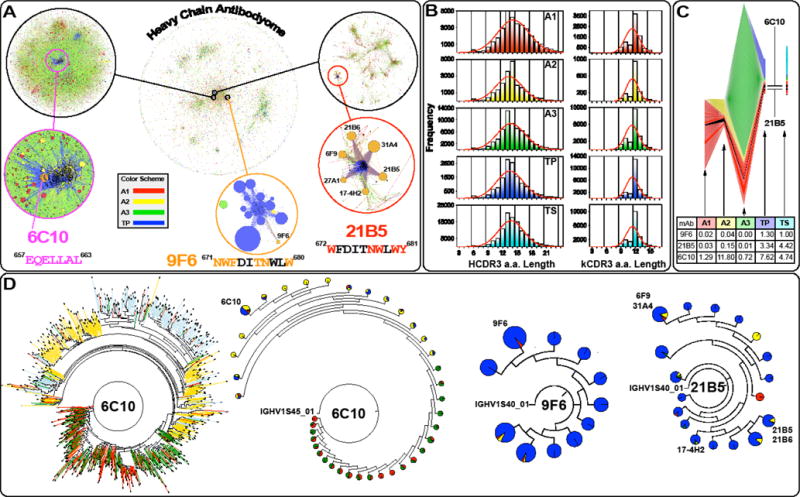
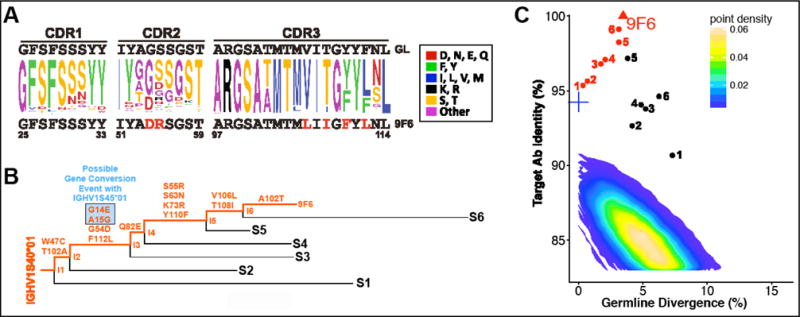
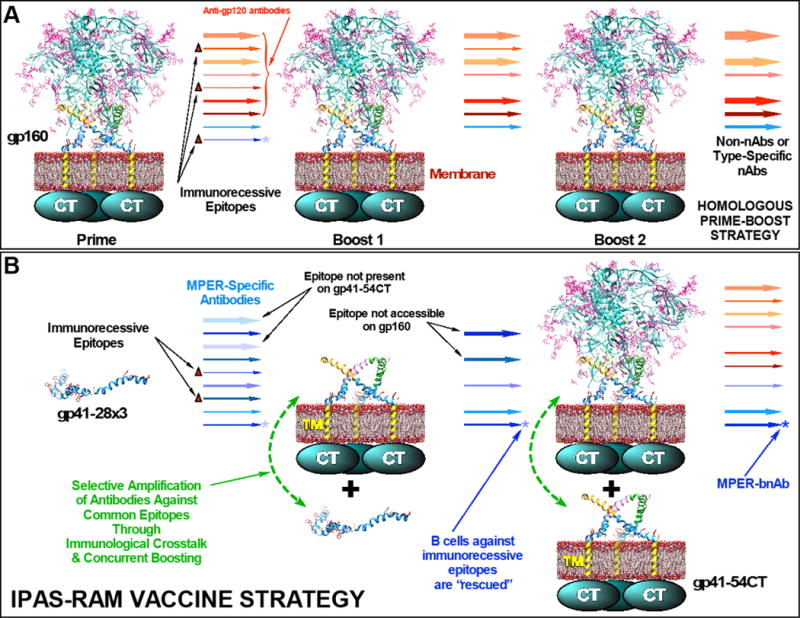
The membrane proximal external region (MPER) of HIV-1 gp41 is targeted by broadly neutralizing antibodies (bnAbs) 4E10 and 10E8. In this proof-of-concept study, we evaluated a novel multi-immunogen vaccine strategy referred to as Incremental, Phased Antigenic Stimulation for Rapid Antibody Maturation (IPAS-RAM) to induce 4E10/10E8-like bnAbs. Rabbits were immunized sequentially, but in a phased manner, with three immunogens that are progressively more native (gp41-28×3, gp41-54CT, and rVV-gp160DH12). Although nAbs were not induced, epitope-mapping analyses indicated that IPAS-RAM vaccination was better able to target antibodies towards the 4E10/10E8 epitopes than homologous prime-boost immunization using gp41-28×3 alone. MPER-specific rabbit monoclonal antibodies were generated, including 9F6. Although it lacked neutralizing activity, the target epitope profile of 9F6 closely resembled those of 4E10 and 10E8 (671NWFDITNWLWYIK683). B-cell repertoire analyses suggested the importance of co-immunizations for maturation of 9F6, which warrants further evaluation of our IPAS-RAM vaccine strategy using an improved priming immunogen.
Keywords: Antibody maturation; HIV-1; MPER; NGS; Neutralizing antibody; Next-generation sequencing; Rabbit; Vaccine; gp41.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Immunogenic properties of a trimeric gp41-based immunogen containing an exposed membrane-proximal external region.Virology. 2015 Dec;486:187-97. doi: 10.1016/j.virol.2015.09.010. Epub 2015 Oct 9. Virology. 2015. PMID: 26454663 Free PMC article.
-
Exposure of the HIV-1 broadly neutralizing antibody 10E8 MPER epitope on the membrane surface by gp41 transmembrane domain scaffolds.Biochim Biophys Acta Biomembr. 2018 Jun;1860(6):1259-1271. doi: 10.1016/j.bbamem.2018.02.019. Epub 2018 Feb 23. Biochim Biophys Acta Biomembr. 2018. PMID: 29477358
-
N-terminal residues of an HIV-1 gp41 membrane-proximal external region antigen influence broadly neutralizing 2F5-like antibodies.Virol Sin. 2015 Dec;30(6):449-56. doi: 10.1007/s12250-015-3664-6. Epub 2015 Dec 21. Virol Sin. 2015. PMID: 26715302 Free PMC article.
-
Neutralizing Antibodies Targeting HIV-1 gp41.Viruses. 2020 Oct 23;12(11):1210. doi: 10.3390/v12111210. Viruses. 2020. PMID: 33114242 Free PMC article. Review.
-
Antigp41 membrane proximal external region antibodies and the art of using the membrane for neutralization.Curr Opin HIV AIDS. 2017 May;12(3):250-256. doi: 10.1097/COH.0000000000000364. Curr Opin HIV AIDS. 2017. PMID: 28422789 Review.
Cited by
-
Brief introduction of current technologies in isolation of broadly neutralizing HIV-1 antibodies.Virus Res. 2018 Jan 2;243:75-82. doi: 10.1016/j.virusres.2017.10.011. Epub 2017 Oct 16. Virus Res. 2018. PMID: 29051051 Free PMC article. Review.
-
Identification of variant HIV envelope proteins with enhanced affinities for precursors to anti-gp41 broadly neutralizing antibodies.PLoS One. 2019 Sep 10;14(9):e0221550. doi: 10.1371/journal.pone.0221550. eCollection 2019. PLoS One. 2019. PMID: 31504041 Free PMC article.
-
Resolving the Molecular Steps in Clostridial Neurotoxin Light Chain Translocation.J Exp Neurol. 2020;1(4):123-134. doi: 10.33696/Neurol.1.020. J Exp Neurol. 2020. PMID: 33615314 Free PMC article.
-
Tracing Antibody Repertoire Evolution by Systems Phylogeny.Front Immunol. 2018 Oct 2;9:2149. doi: 10.3389/fimmu.2018.02149. eCollection 2018. Front Immunol. 2018. PMID: 30333820 Free PMC article. Review.
-
Automated analysis of immunosequencing datasets reveals novel immunoglobulin D genes across diverse species.PLoS Comput Biol. 2020 Apr 27;16(4):e1007837. doi: 10.1371/journal.pcbi.1007837. eCollection 2020 Apr. PLoS Comput Biol. 2020. PMID: 32339161 Free PMC article.
References
-
- Appella E, Chersi A, Rejnek J, Reisfeld R, Mage R. Rabbit immunoglobulin lambda chains: isolation and amino acid sequence of cysteine-containing peptides. Immunochemistry. 1974;11:395–402. - PubMed
-
- Bastian M, Heymann S, Jacomy M. Gephi: an open source software for exploring and manipulating networks. ICWSM 2009
-
- Briney B, Sok D, Jardine JG, Kulp DW, Skog P, Menis S, Jacak R, Kalyuzhniy O, de Val N, Sesterhenn F, Le KM, Ramos A, Jones M, Saye-Francisco KL, Blane TR, Spencer S, Georgeson E, Hu X, Ozorowski G, Adachi Y, Kubitz M, Sarkar A, Wilson IA, Ward AB, Nemazee D, Burton DR, Schief WR. Tailored Immunogens Direct Affinity Maturation toward HIV Neutralizing Antibodies. Cell. 2016;166:1459–1470.e11. doi: 10.1016/j.cell.2016.08.005. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
