

Diverse Non-genetic, Allele-Specific Expression Effects Shape Genetic Architecture at the Cellular Level in the Mammalian Brain
- PMID: 28238550
- PMCID: PMC5774018
- DOI: 10.1016/j.neuron.2017.01.033
Diverse Non-genetic, Allele-Specific Expression Effects Shape Genetic Architecture at the Cellular Level in the Mammalian Brain
Abstract
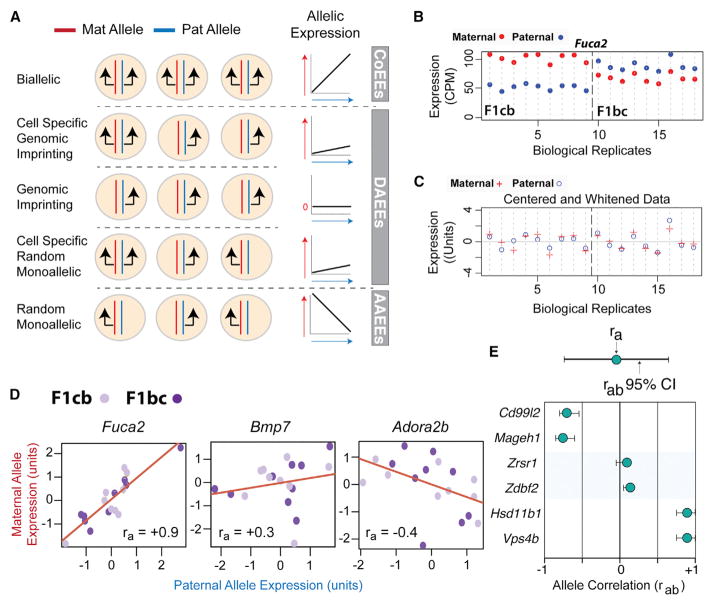
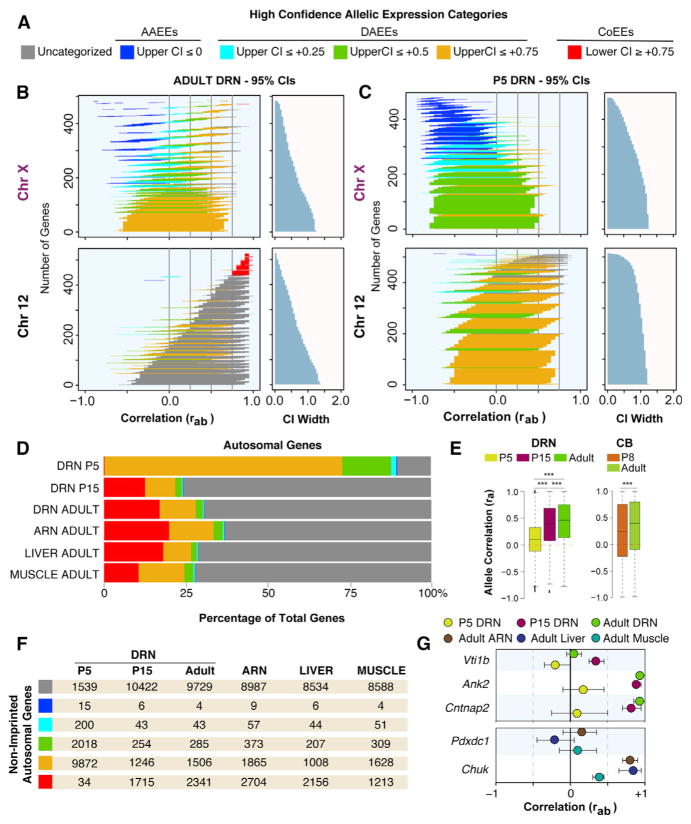
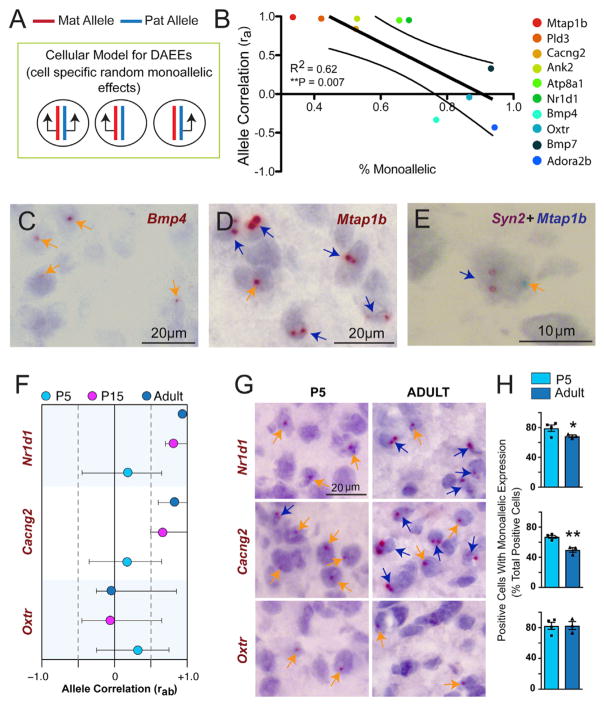
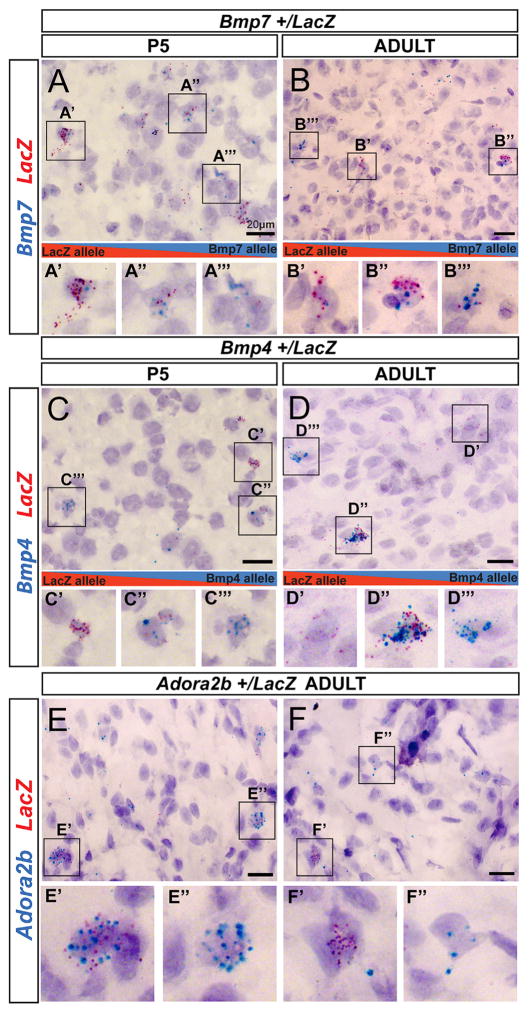
Interactions between genetic and epigenetic effects shape brain function, behavior, and the risk for mental illness. Random X inactivation and genomic imprinting are epigenetic allelic effects that are well known to influence genetic architecture and disease risk. Less is known about the nature, prevalence, and conservation of other potential epigenetic allelic effects in vivo in the mouse and primate brain. Here we devise genomics, in situ hybridization, and mouse genetics strategies to uncover diverse allelic effects in the brain that are not caused by imprinting or genetic variation. We found allelic effects that are developmental stage and cell type specific, that are prevalent in the neonatal brain, and that cause mosaics of monoallelic brain cells that differentially express wild-type and mutant alleles for heterozygous mutations. Finally, we show that diverse non-genetic allelic effects that impact mental illness risk genes exist in the macaque and human brain. Our findings have potential implications for mammalian brain genetics. VIDEO ABSTRACT.
Keywords: allele-specific expression; autism; behavioral genetics; brain development; brain epigenetics; genomic imprinting; mental illness; monoallelic; random monoallelic expression; transcriptome.
Published by Elsevier Inc.
Figures
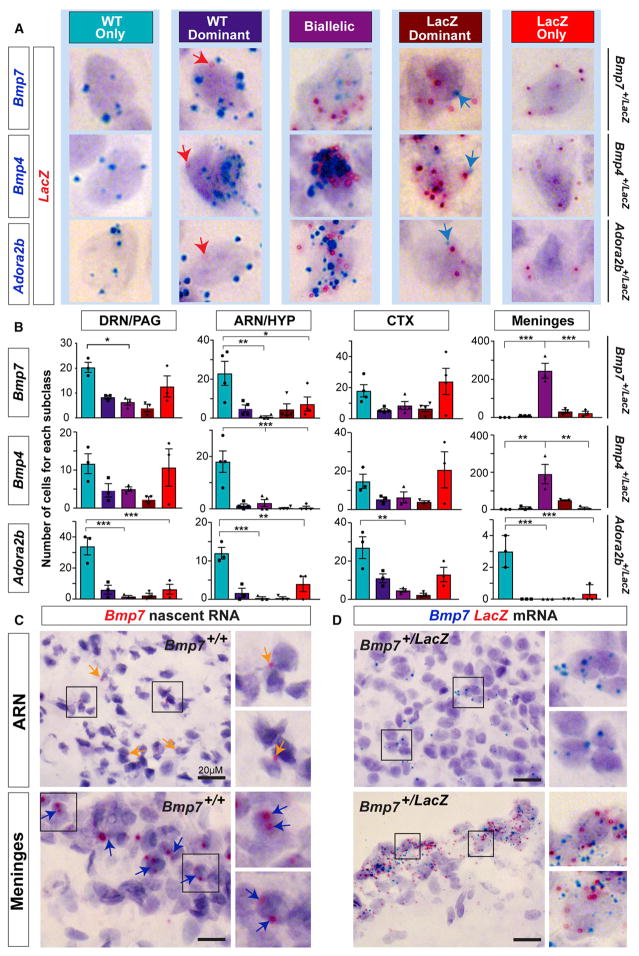
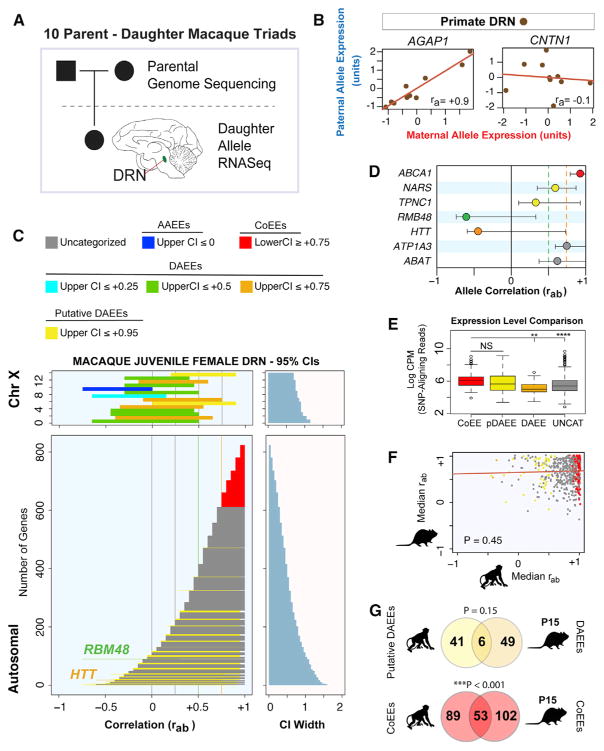
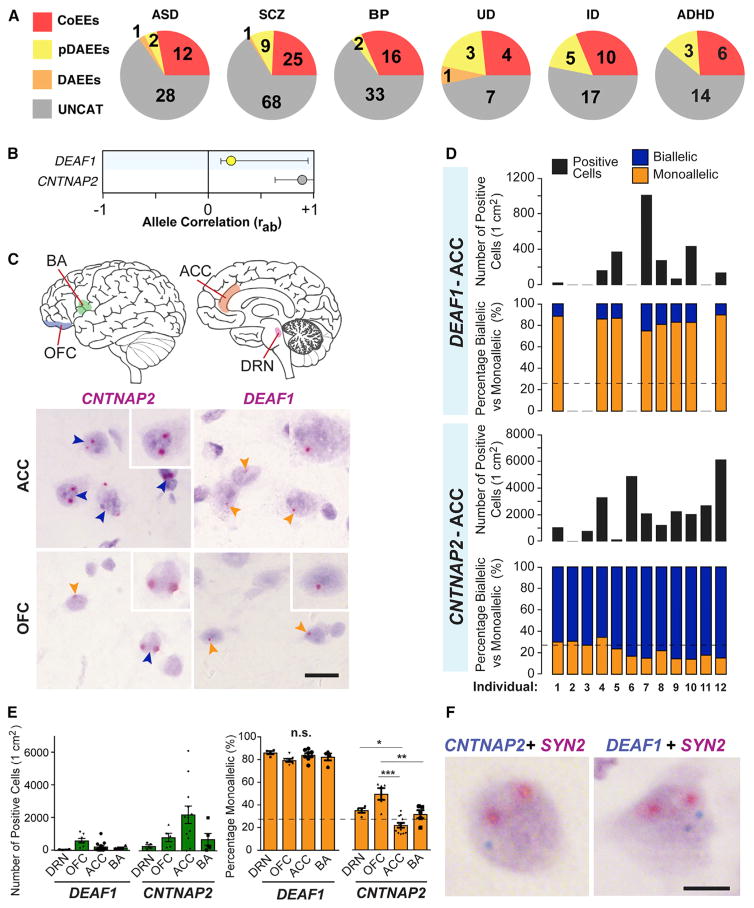
References
-
- Amaral DG, Schumann CM, Nordahl CW. Neuroanatomy of autism. Trends Neurosci. 2008;31:137–145. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
