

RTN3 Is a Novel Cold-Induced Protein and Mediates Neuroprotective Effects of RBM3
- PMID: 28238655
- PMCID: PMC5344685
- DOI: 10.1016/j.cub.2017.01.047
RTN3 Is a Novel Cold-Induced Protein and Mediates Neuroprotective Effects of RBM3
Abstract
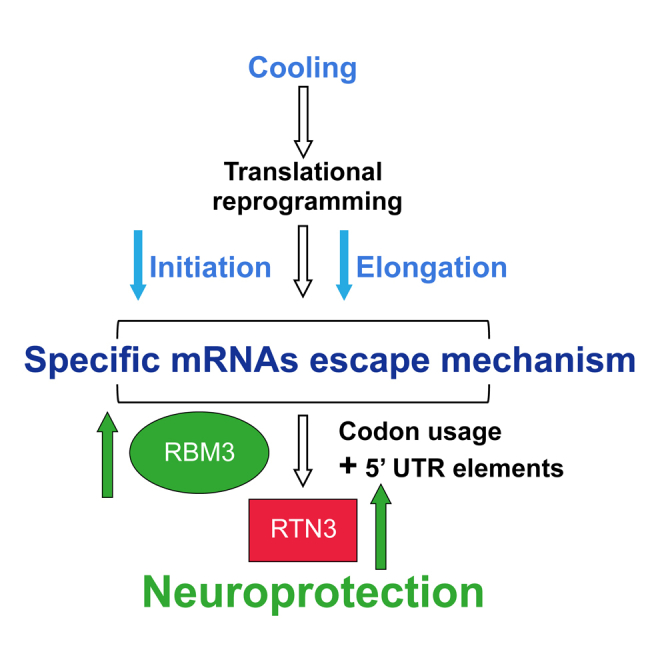
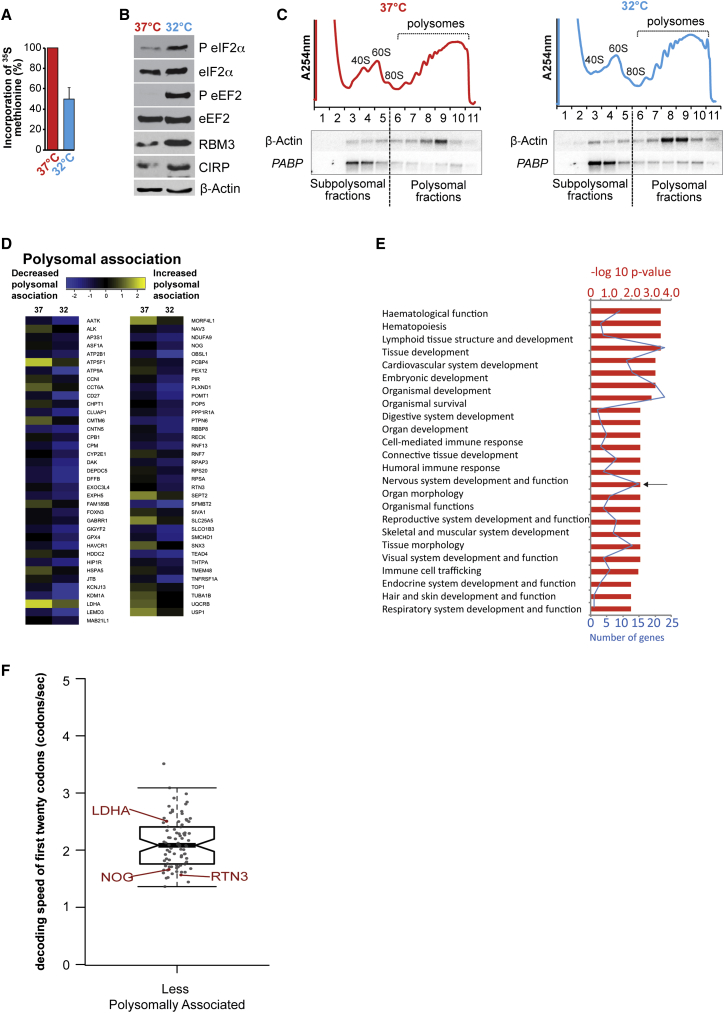
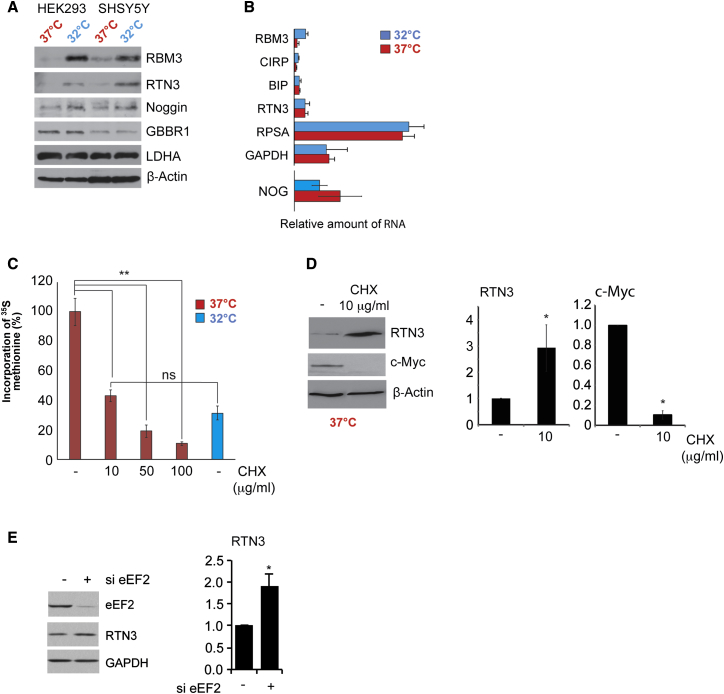
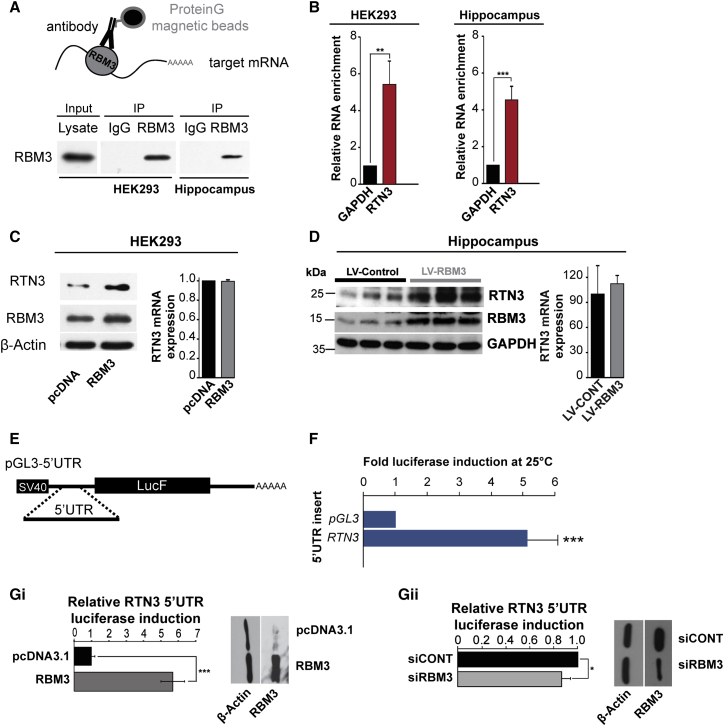
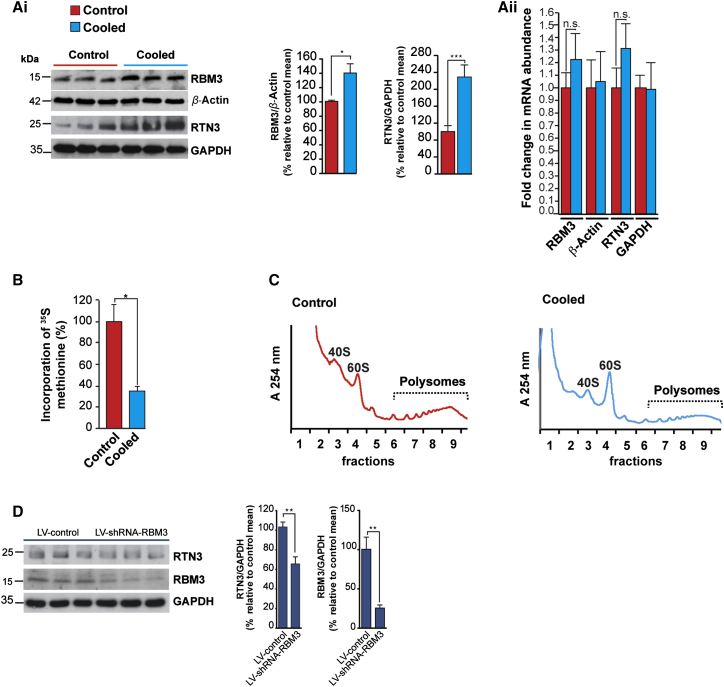
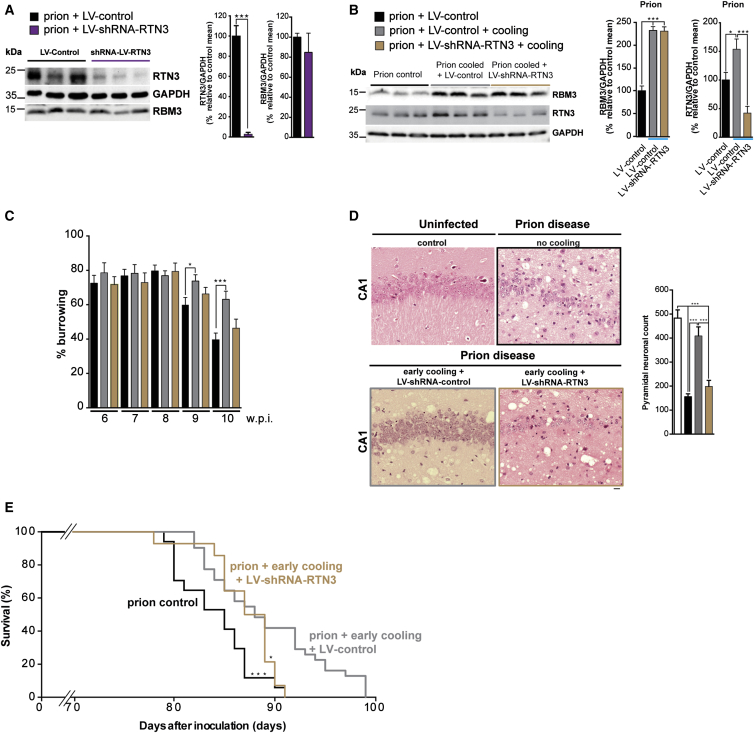
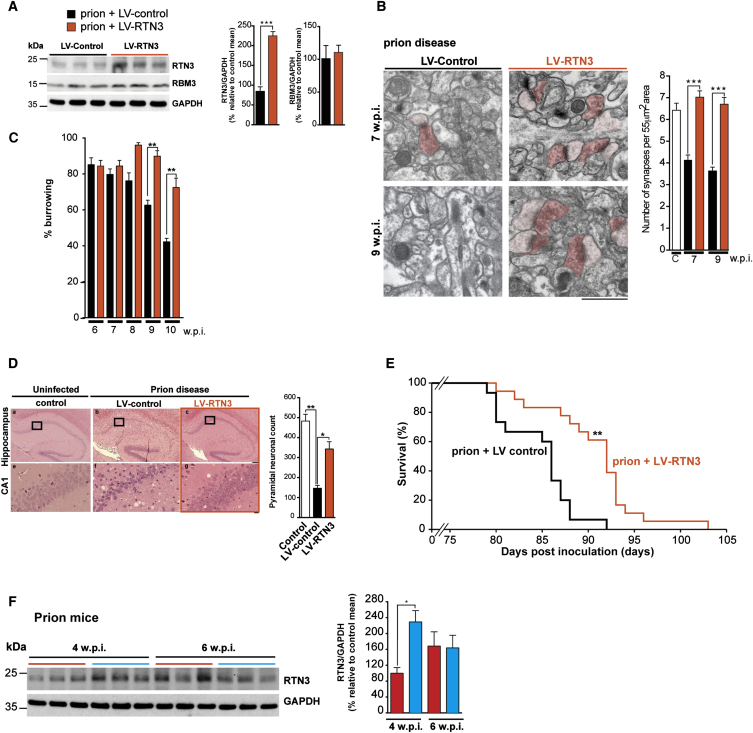
Cooling and hypothermia are profoundly neuroprotective, mediated, at least in part, by the cold shock protein, RBM3. However, the neuroprotective effector proteins induced by RBM3 and the mechanisms by which mRNAs encoding cold shock proteins escape cooling-induced translational repression are unknown. Here, we show that cooling induces reprogramming of the translatome, including the upregulation of a new cold shock protein, RTN3, a reticulon protein implicated in synapse formation. We report that this has two mechanistic components. Thus, RTN3 both evades cooling-induced translational elongation repression and is also bound by RBM3, which drives the increased expression of RTN3. In mice, knockdown of RTN3 expression eliminated cooling-induced neuroprotection. However, lentivirally mediated RTN3 overexpression prevented synaptic loss and cognitive deficits in a mouse model of neurodegeneration, downstream and independently of RBM3. We conclude that RTN3 expression is a mediator of RBM3-induced neuroprotection, controlled by novel mechanisms of escape from translational inhibition on cooling.
Keywords: RBM3; RTN3; cold shock; mRNA translation; neuroprotection; protein synthesis.
Copyright © 2017 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Figures
Comment in
-
Hypothermia-induced neuroprotection.Sci Signal. 2017 Mar 14;10(470):eaan1410. doi: 10.1126/scisignal.aan1410. Sci Signal. 2017. PMID: 28292952
References
-
- Delhaye C., Mahmoudi M., Waksman R. Hypothermia therapy: neurological and cardiac benefits. J. Am. Coll. Cardiol. 2012;59:197–210. - PubMed
-
- Yenari M.A., Han H.S. Neuroprotective mechanisms of hypothermia in brain ischaemia. Nat. Rev. Neurosci. 2012;13:267–278. - PubMed
-
- Wellmann S., Bührer C., Moderegger E., Zelmer A., Kirschner R., Koehne P., Fujita J., Seeger K. Oxygen-regulated expression of the RNA-binding proteins RBM3 and CIRP by a HIF-1-independent mechanism. J. Cell Sci. 2004;117:1785–1794. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
