

Functional and Structural Analysis of Programmed C-Methylation in the Biosynthesis of the Fungal Polyketide Citrinin
- PMID: 28238725
- PMCID: PMC5419425
- DOI: 10.1016/j.chembiol.2017.01.008
Functional and Structural Analysis of Programmed C-Methylation in the Biosynthesis of the Fungal Polyketide Citrinin
Abstract
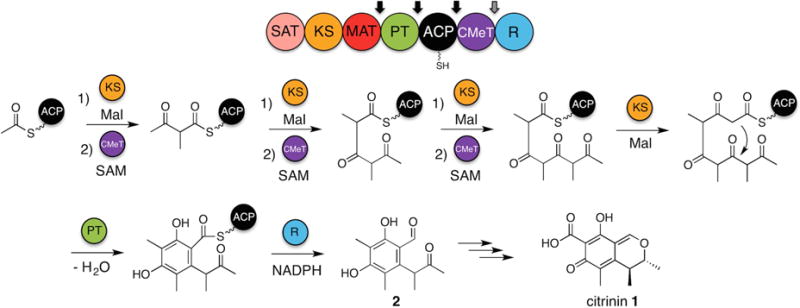
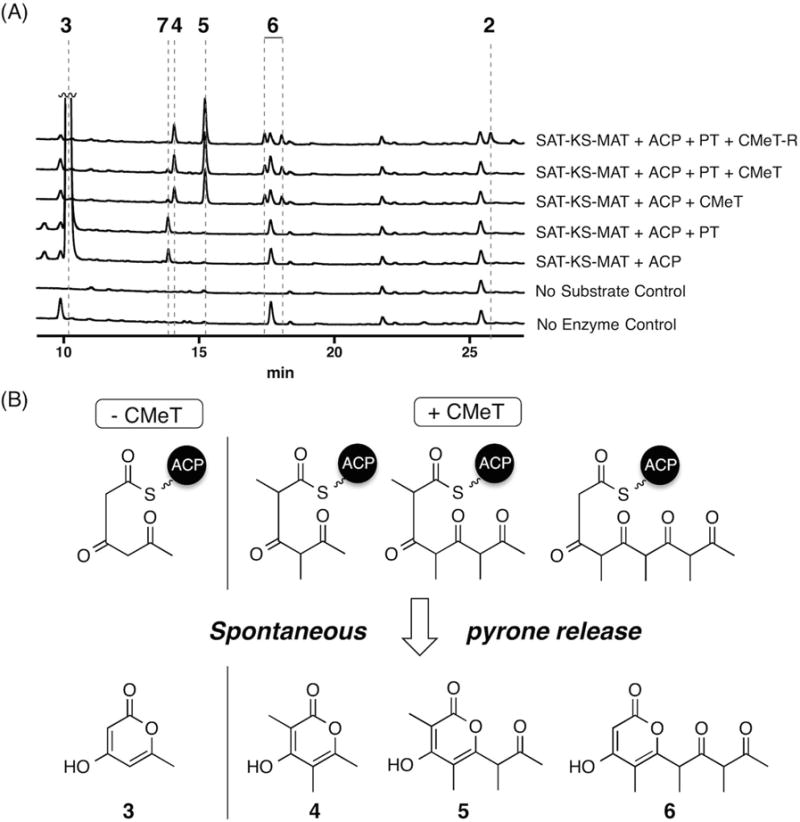
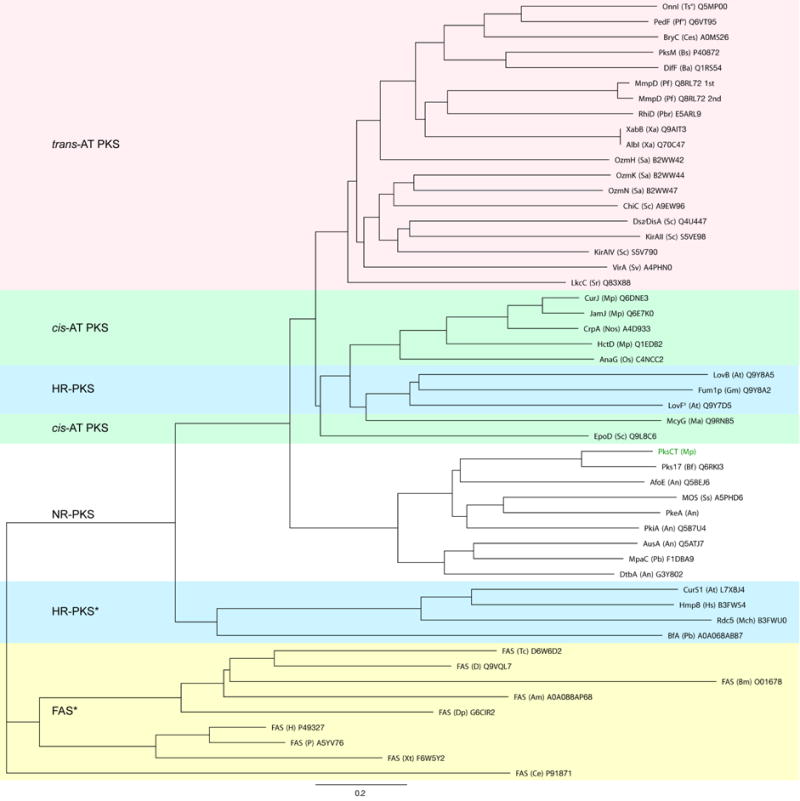
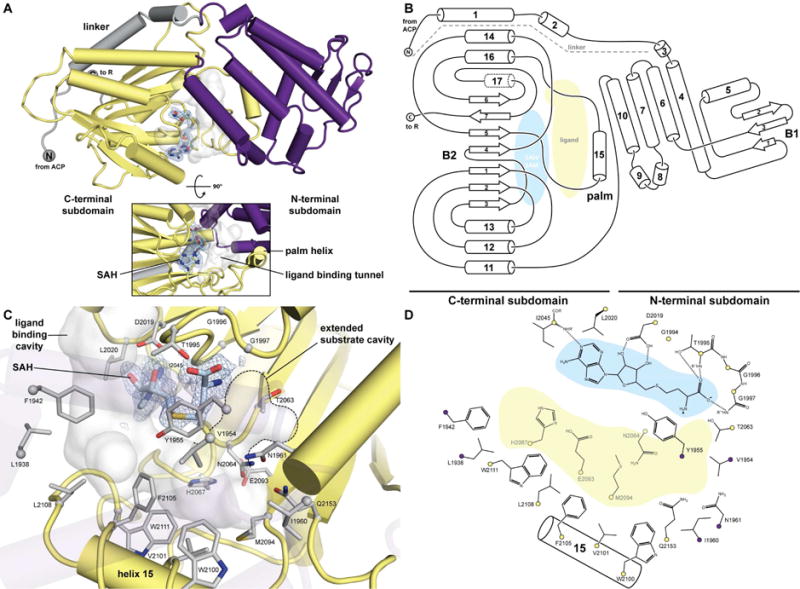
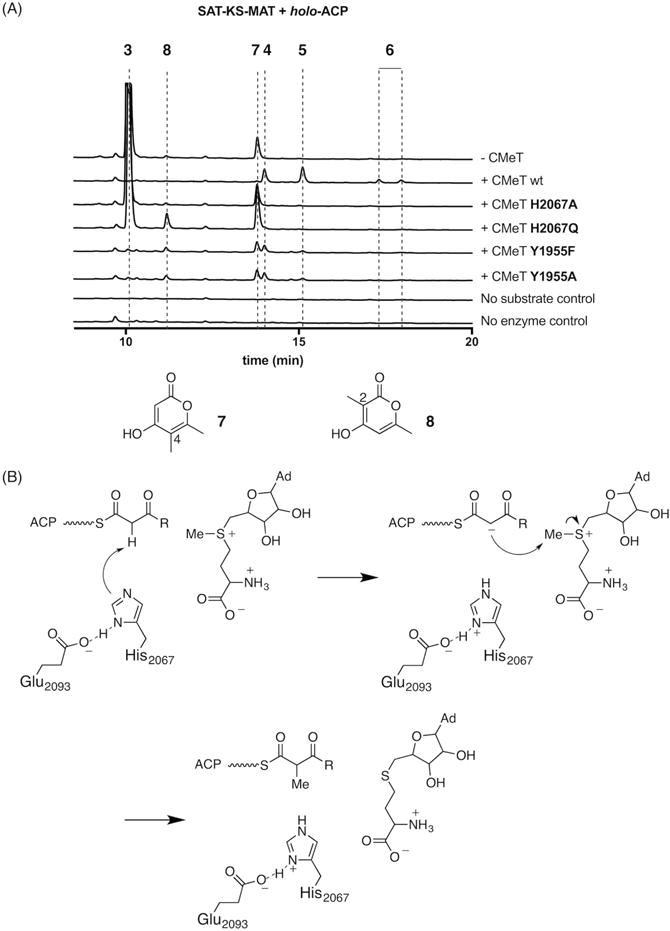
Fungal polyketide synthases (PKSs) are large, multidomain enzymes that biosynthesize a wide range of natural products. A hallmark of these megasynthases is the iterative use of catalytic domains to extend and modify a series of enzyme-bound intermediates. A subset of these iterative PKSs (iPKSs) contains a C-methyltransferase (CMeT) domain that adds one or more S-adenosylmethionine (SAM)-derived methyl groups to the carbon framework. Neither the basis by which only specific positions on the growing intermediate are methylated ("programming") nor the mechanism of methylation are well understood. Domain dissection and reconstitution of PksCT, the fungal non-reducing PKS (NR-PKS) responsible for the first isolable intermediate in citrinin biosynthesis, demonstrates the role of CMeT-catalyzed methylation in precursor elongation and pentaketide formation. The crystal structure of the S-adenosyl-homocysteine (SAH) coproduct-bound PksCT CMeT domain reveals a two-subdomain organization with a novel N-terminal subdomain characteristic of PKS CMeT domains and provides insights into co-factor and ligand recognition.
Keywords: C-methylation; PKS; catalytic programming; citrinin; domain deconstruction; iterative biosynthesis; polyketide; structure.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- Cacho RA, Thuss J, Xu W, Sanichar R, Gao Z, Nguyen A, Vederas JC, Tang Y. Understanding Programming of Fungal Iterative Polyketide Synthases: The Biochemical Basis for Regioselectivity by the Methyltransferase Domain in the Lovastatin Megasynthase. J Am Chem Soc. 2015;137:15688–15691. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
