

A laboratory simulation of Arabidopsis seed dormancy cycling provides new insight into its regulation by clock genes and the dormancy-related genes DOG1, MFT, CIPK23 and PHYA
- PMID: 28240777
- PMCID: PMC5518234
- DOI: 10.1111/pce.12940
A laboratory simulation of Arabidopsis seed dormancy cycling provides new insight into its regulation by clock genes and the dormancy-related genes DOG1, MFT, CIPK23 and PHYA
Abstract
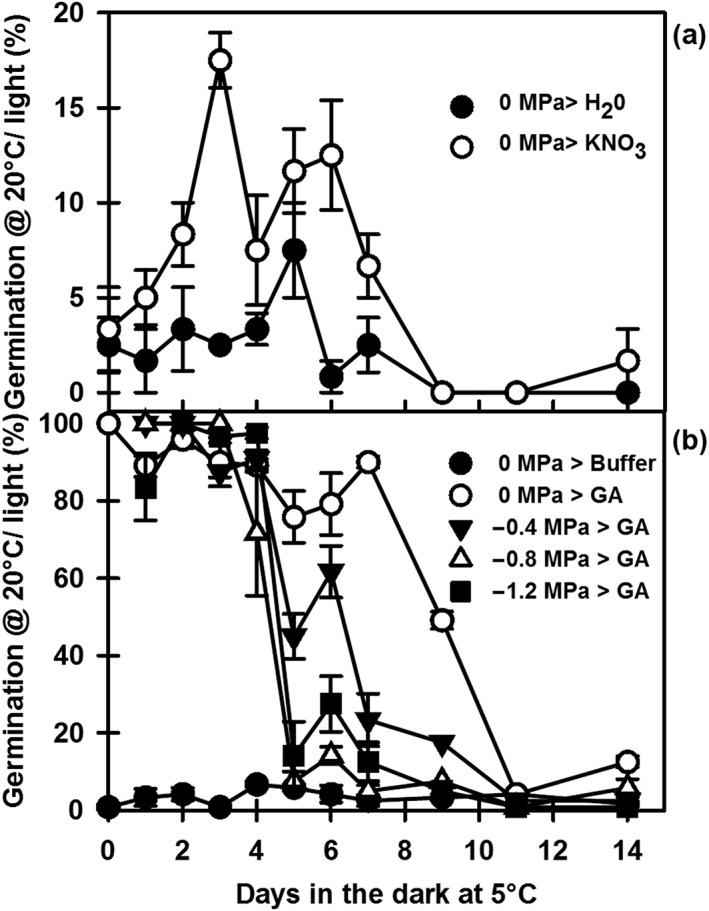
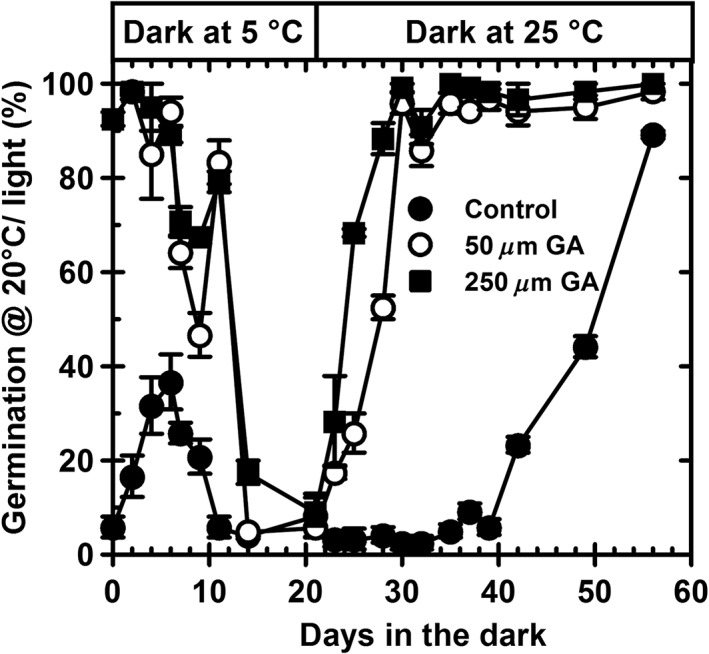
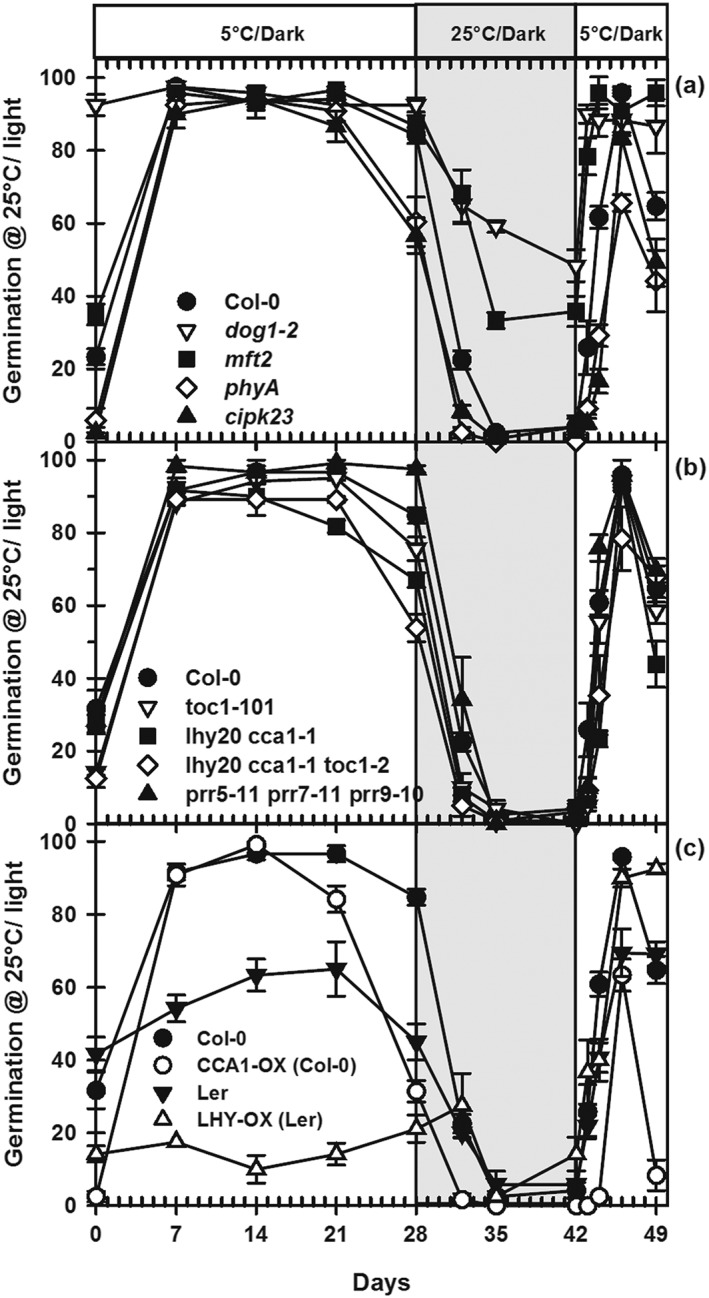
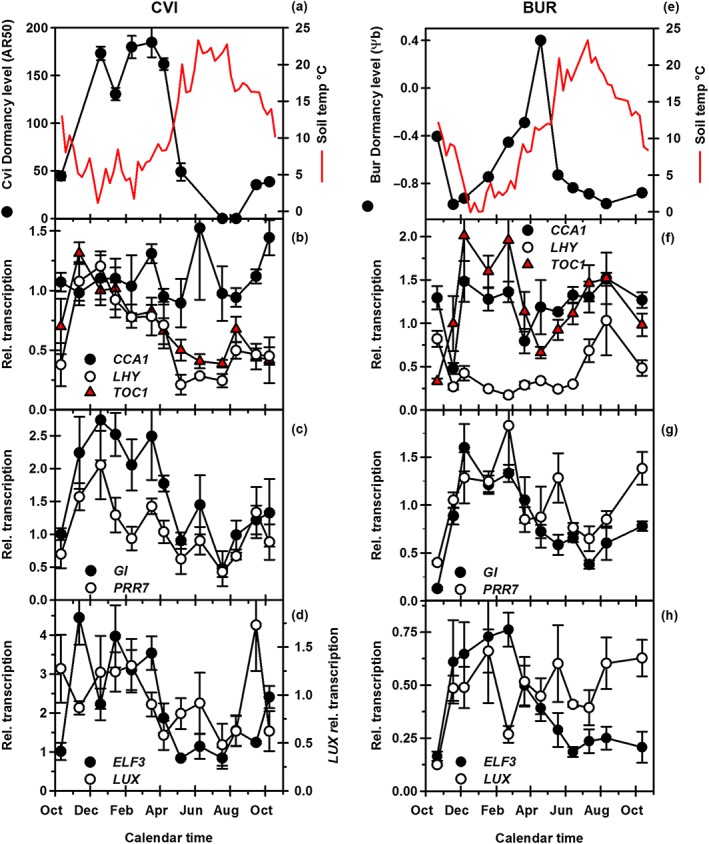
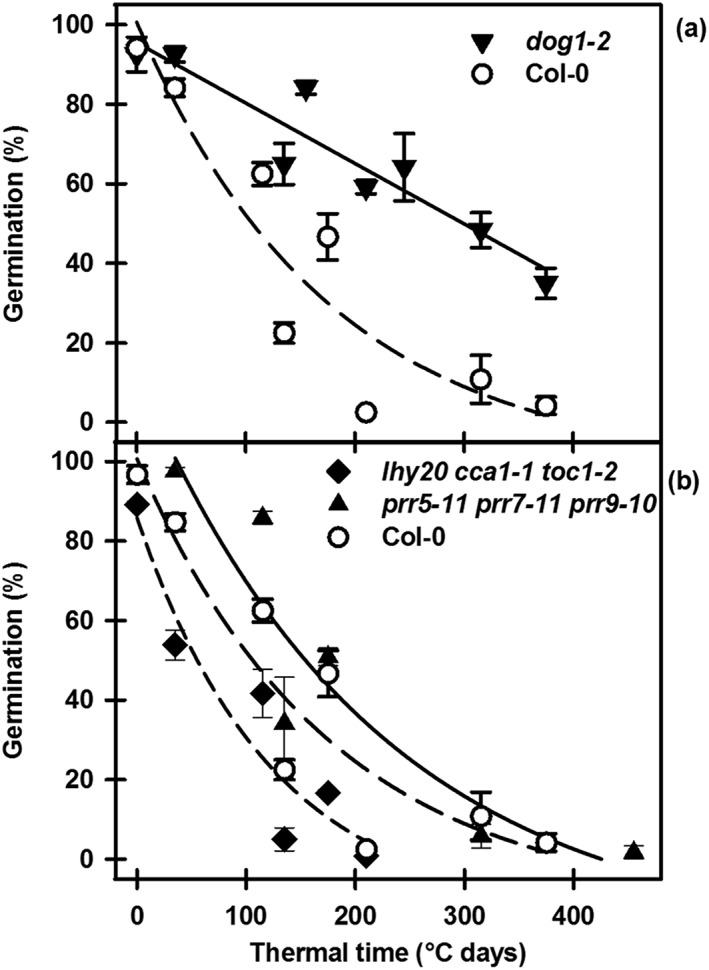
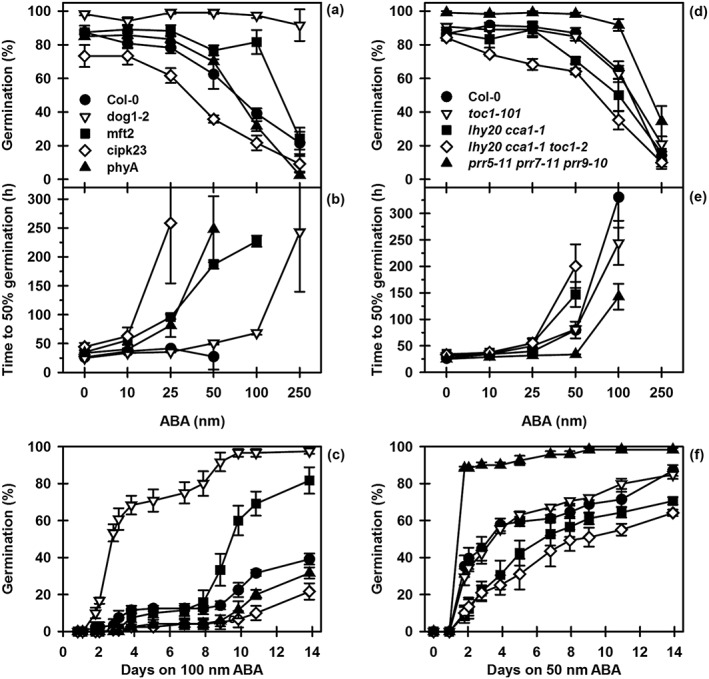
Environmental signals drive seed dormancy cycling in the soil to synchronize germination with the optimal time of year, a process essential for species' fitness and survival. Previous correlation of transcription profiles in exhumed seeds with annual environmental signals revealed the coordination of dormancy-regulating mechanisms with the soil environment. Here, we developed a rapid and robust laboratory dormancy cycling simulation. The utility of this simulation was tested in two ways: firstly, using mutants in known dormancy-related genes [DELAY OF GERMINATION 1 (DOG1), MOTHER OF FLOWERING TIME (MFT), CBL-INTERACTING PROTEIN KINASE 23 (CIPK23) and PHYTOCHROME A (PHYA)] and secondly, using further mutants, we test the hypothesis that components of the circadian clock are involved in coordination of the annual seed dormancy cycle. The rate of dormancy induction and relief differed in all lines tested. In the mutants, dog1-2 and mft2, dormancy induction was reduced but not absent. DOG1 is not absolutely required for dormancy. In cipk23 and phyA dormancy, induction was accelerated. Involvement of the clock in dormancy cycling was clear when mutants in the morning and evening loops of the clock were compared. Dormancy induction was faster when the morning loop was compromised and delayed when the evening loop was compromised.
Keywords: circadian clock; circannual rhythm; germination; thermal time.
© 2017 The Authors Plant, Cell & Environment Published by John Wiley & Sons Ltd.
Figures
References
-
- Atamian H.S. & Harmer S.L. (2016) Circadian regulation of hormone signaling and plant physiology. Plant Molecular Biology 91, 691–702. - PubMed
-
- Auge G.A., Blair L.K., Burghardt L.T., Coughlan J., Edwards B., Leverett L.D. & Donohue K. (2015) Secondary dormancy dynamics depends on primary dormancy status in <styled-content style="fixed-case"><styled-content style="italic-in-any-context">Arabidopsis thaliana</styled-content></styled-content> . Seed Science Research 25, 230–246.
-
- Baskin C.C. & Baskin J.M. (1998) Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. Academic Press, San Diego.
-
- Batlla D., Grundy A., Dent K.C., Clay H.A. & Finch‐Savage W.E. (2009) A quantitative analysis of temperature‐dependent dormancy changes in <styled-content style="fixed-case"><styled-content style="italic-in-any-context">Polygonum aviculare</styled-content></styled-content> seeds. Weed Research 49, 428–438.
-
- Batlla D. & Benech‐Arnold R.L. (2014) Weed seed germination and the light environment: implications for weed management. Weed Biology and Management 14, 77–87.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
