

Malaria Parasite Liver Infection and Exoerythrocytic Biology
- PMID: 28242785
- PMCID: PMC5453383
- DOI: 10.1101/cshperspect.a025486
Malaria Parasite Liver Infection and Exoerythrocytic Biology
Abstract
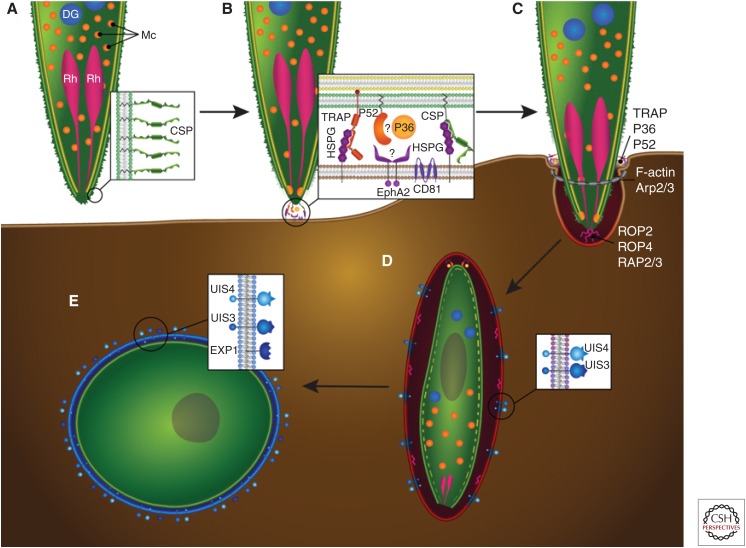
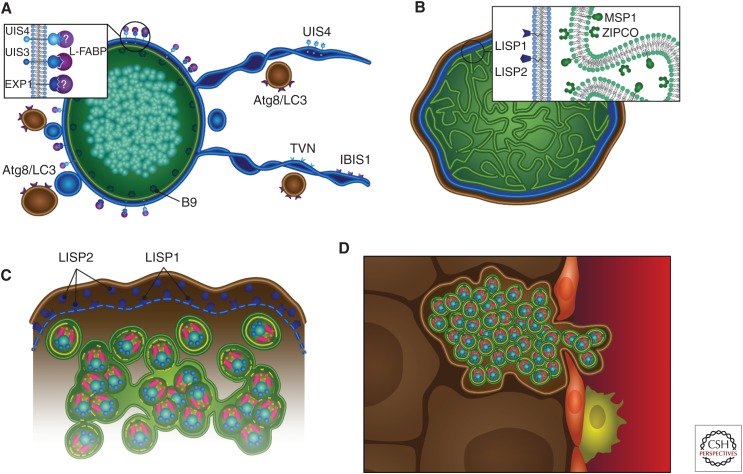
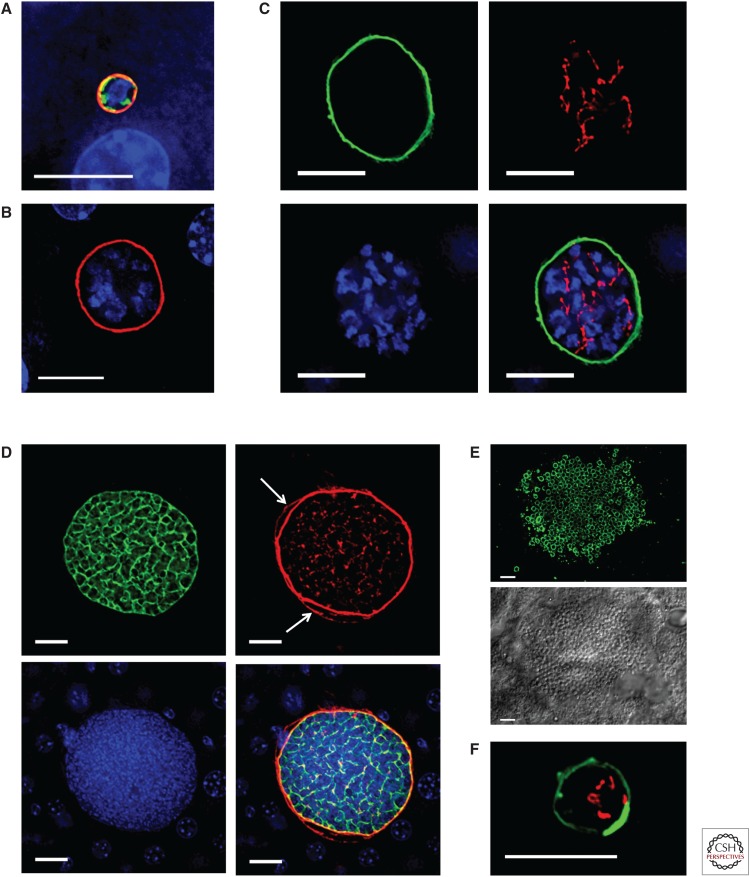
In their infection cycle, malaria parasites undergo replication and population expansions within the vertebrate host and the mosquito vector. Host infection initiates with sporozoite invasion of hepatocytes, followed by a dramatic parasite amplification event during liver stage parasite growth and replication within hepatocytes. Each liver stage forms up to 90,000 exoerythrocytic merozoites, which are in turn capable of initiating a blood stage infection. Liver stages not only exploit host hepatocyte resources for nutritional needs but also endeavor to prevent hepatocyte cell death and detection by the host's immune system. Research over the past decade has identified numerous parasite factors that play a critical role during liver infection and has started to delineate a complex web of parasite-host interactions that sustain successful parasite colonization of the mammalian host. Targeting the parasites' obligatory infection of the liver as a gateway to the blood, with drugs and vaccines, constitutes the most effective strategy for malaria eradication, as it would prevent clinical disease and onward transmission of the parasite.
Copyright © 2017 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Annoura T, van Schaijk BC, Ploemen IH, Sajid M, Lin JW, Vos MW, Dinmohamed AG, Inaoka DK, Rijpma SR, van Gemert GJ, et al. 2014. Two Plasmodium 6-Cys family-related proteins have distinct and critical roles in liver-stage development. FASEB J 28: 2158–2170. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous