

Critical Role of Myeloid-Derived Suppressor Cells in Tumor-Induced Liver Immune Suppression through Inhibition of NKT Cell Function
- PMID: 28243237
- PMCID: PMC5303828
- DOI: 10.3389/fimmu.2017.00129
Critical Role of Myeloid-Derived Suppressor Cells in Tumor-Induced Liver Immune Suppression through Inhibition of NKT Cell Function
Abstract
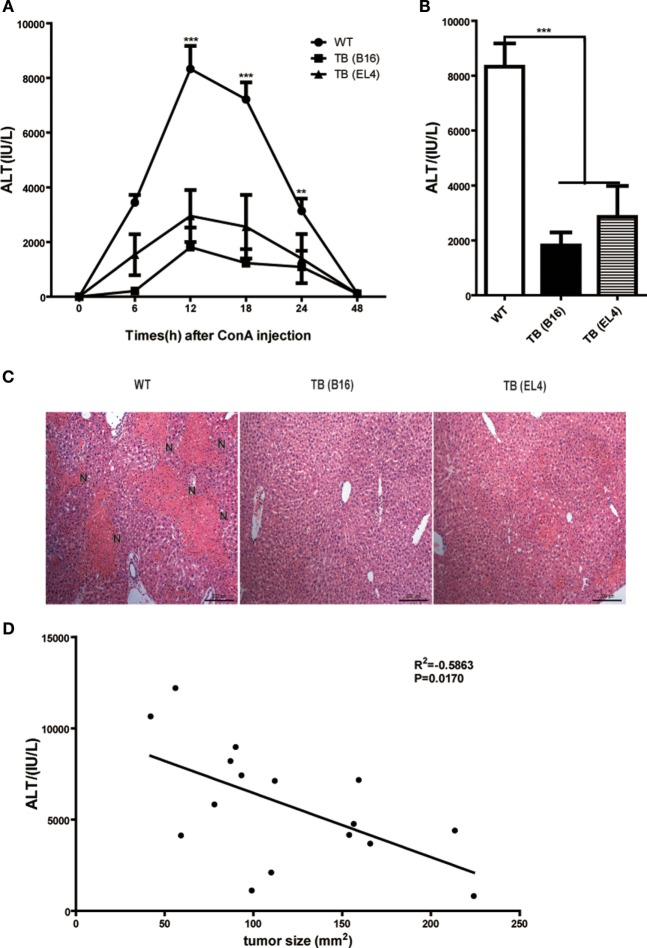
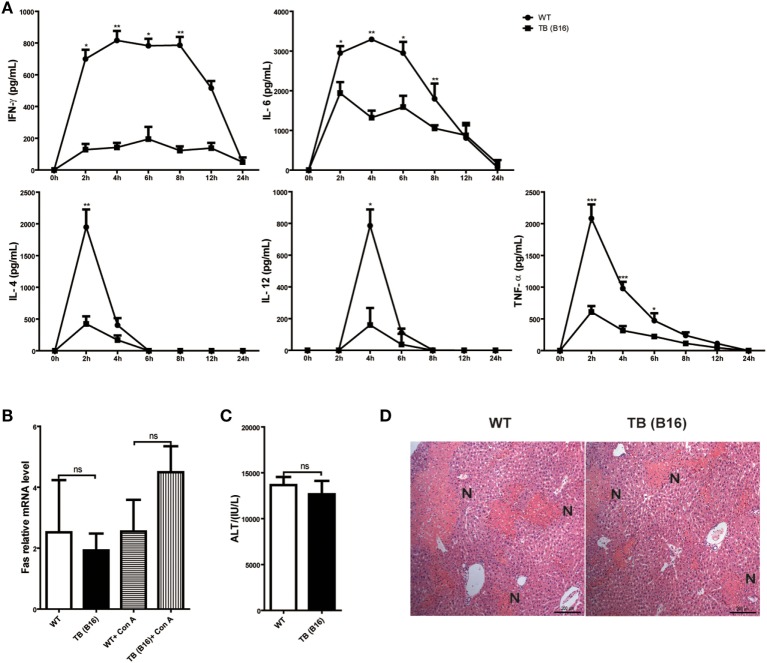
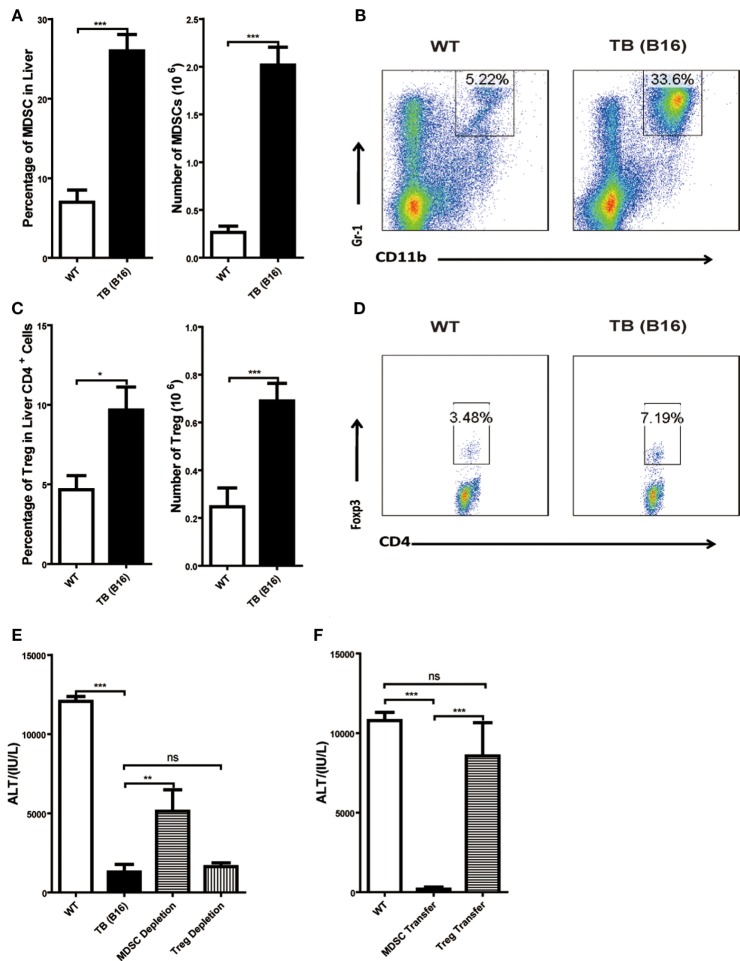
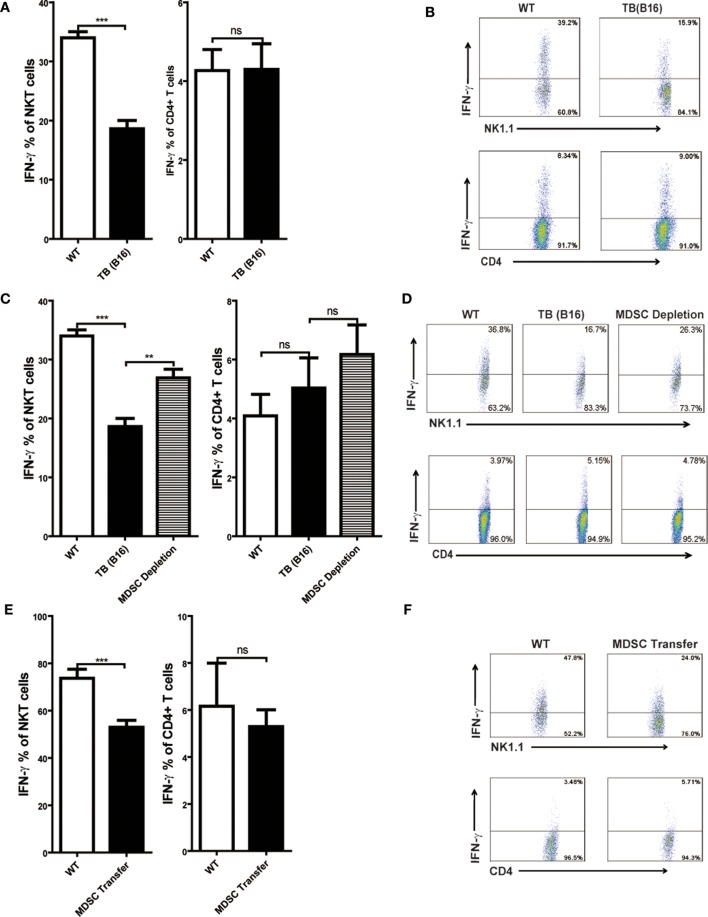
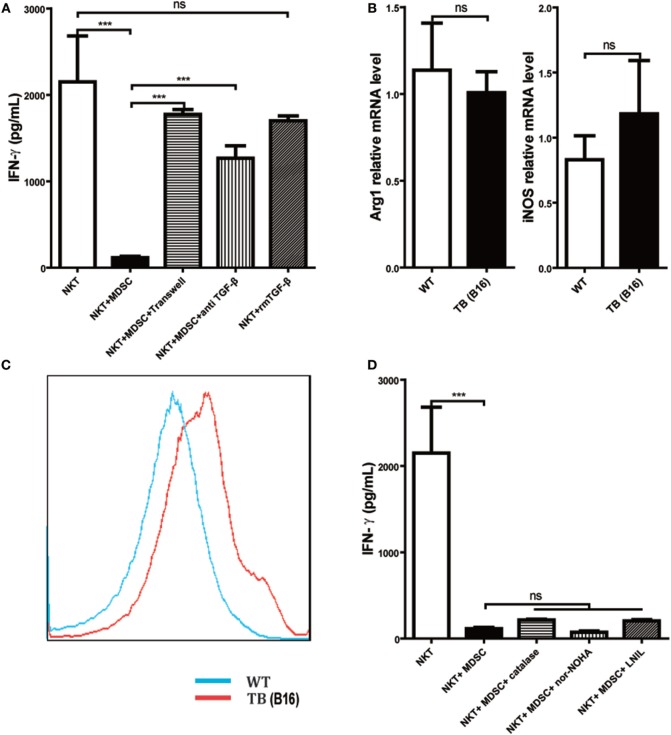
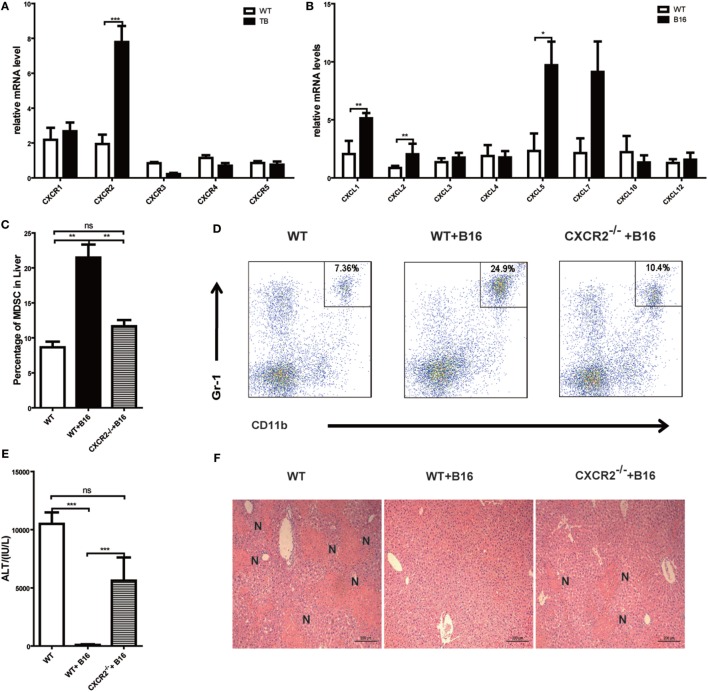
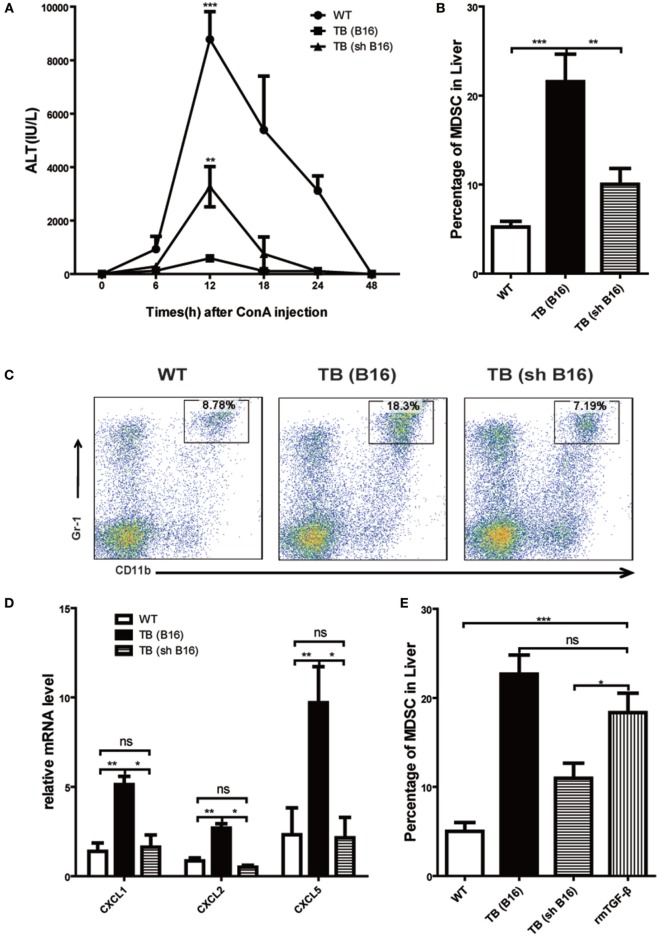
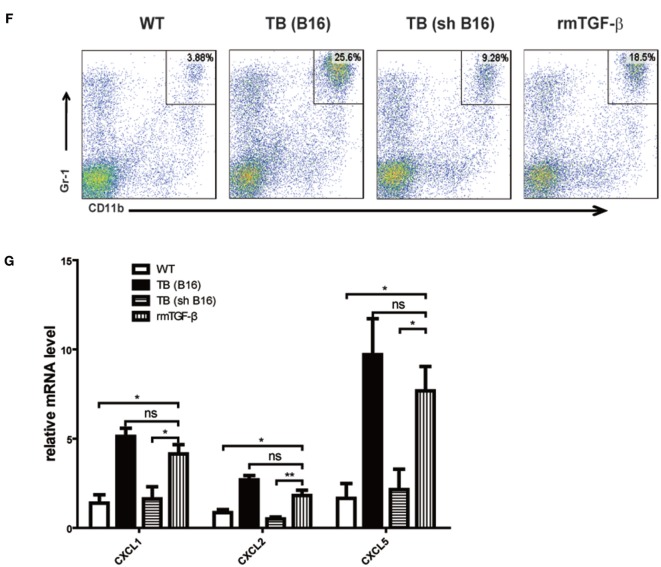
Metastasis followed by the tumor development is the primary cause of death for cancer patients. However, the underlying molecular mechanisms of how the growth of tumor resulted in the immune suppression, especially at the blood-enriched organ such as liver, were largely unknown. In this report, we studied the liver immune response of tumor-bearing (TB) mice using concanavalin A (Con A)-induced hepatitis model. We demonstrated that TB mice displayed an immune suppression phenotype, with attenuated alanine aminotransferase levels and liver damage upon Con A treatment. We also elucidated that large amounts of myeloid-derived suppressor cells (MDSCs) being influx into the liver in TB mice and these MDSCs were essential for liver immune suppression through both depletion and reconstitution approaches. We further determined that these MDSCs selectively suppressed the IFN-γ production deriving from NKT cells through membrane-bound transforming growth factor β (TGF-β). Finally, we defined a tumor-derived TGF-β-triggered CXCL1/2/5- and CXCR2-dependent recruitment of MDSC into the liver. In summary, our results defined a novel mechanism of liver immune suppression triggered by growing living tumor and provided possible therapeutic targets against these MDSCs.
Keywords: CXCR2; MDSC; TGF-β; liver immune suppression; remote tumor.
Figures
Similar articles
-
Dopamine inhibits the function of Gr-1+CD115+ myeloid-derived suppressor cells through D1-like receptors and enhances anti-tumor immunity.J Leukoc Biol. 2015 Jan;97(1):191-200. doi: 10.1189/jlb.5A1113-626RR. Epub 2014 Oct 23. J Leukoc Biol. 2015. PMID: 25341727 Free PMC article.
-
SSC(high)CD11b(high)Ly-6C(high)Ly-6G(low) myeloid cells curtail CD4 T cell response by inducible nitric oxide synthase in murine hepatitis.Int J Biochem Cell Biol. 2014 Sep;54:89-97. doi: 10.1016/j.biocel.2014.07.005. Epub 2014 Jul 15. Int J Biochem Cell Biol. 2014. PMID: 25035167
-
Tumor- and organ-dependent infiltration by myeloid-derived suppressor cells.Int Immunopharmacol. 2011 Jul;11(7):816-26. doi: 10.1016/j.intimp.2011.02.021. Epub 2011 Mar 2. Int Immunopharmacol. 2011. PMID: 21376153
-
Myeloid-Derived Suppressor Cells and Therapeutic Strategies in Cancer.Mediators Inflamm. 2015;2015:159269. doi: 10.1155/2015/159269. Epub 2015 May 19. Mediators Inflamm. 2015. PMID: 26078490 Free PMC article. Review.
-
Suppression of T cells by myeloid-derived suppressor cells in cancer.Hum Immunol. 2017 Feb;78(2):113-119. doi: 10.1016/j.humimm.2016.12.001. Epub 2016 Dec 7. Hum Immunol. 2017. PMID: 27939507 Review.
Cited by
-
Myeloid-Derived Suppressor Cells in Tumors: From Mechanisms to Antigen Specificity and Microenvironmental Regulation.Front Immunol. 2020 Jul 22;11:1371. doi: 10.3389/fimmu.2020.01371. eCollection 2020. Front Immunol. 2020. PMID: 32793192 Free PMC article. Review.
-
Bacteroides acidifaciens in the gut plays a protective role against CD95-mediated liver injury.Gut Microbes. 2022 Jan-Dec;14(1):2027853. doi: 10.1080/19490976.2022.2027853. Gut Microbes. 2022. PMID: 35129072 Free PMC article.
-
Regulation of T cells by myeloid-derived suppressor cells: emerging immunosuppressor in lung cancer.Discov Oncol. 2023 Oct 19;14(1):185. doi: 10.1007/s12672-023-00793-1. Discov Oncol. 2023. PMID: 37857728 Free PMC article. Review.
-
Monocytic Myeloid-Derived Suppressor Cells in Chronic Infections.Front Immunol. 2018 Jan 4;8:1895. doi: 10.3389/fimmu.2017.01895. eCollection 2017. Front Immunol. 2018. PMID: 29354120 Free PMC article. Review.
-
Combination of molecularly targeted therapies and immune checkpoint inhibitors in the new era of unresectable hepatocellular carcinoma treatment.Ther Adv Med Oncol. 2021 May 24;13:17588359211018026. doi: 10.1177/17588359211018026. eCollection 2021. Ther Adv Med Oncol. 2021. PMID: 34104226 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources