

Deciphering the BAR code of membrane modulators
- PMID: 28243699
- PMCID: PMC5487894
- DOI: 10.1007/s00018-017-2478-0
Deciphering the BAR code of membrane modulators
Abstract
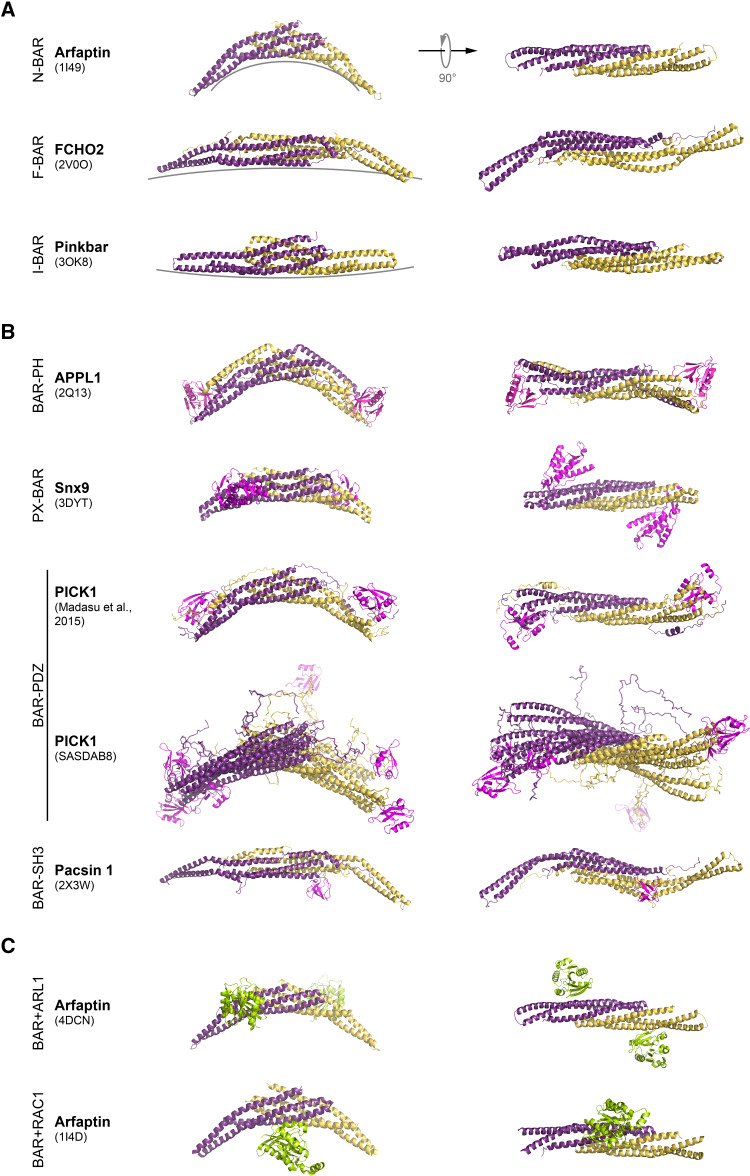
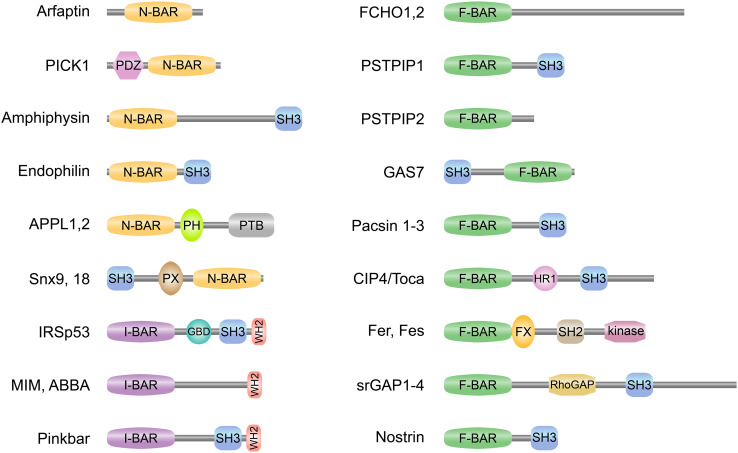
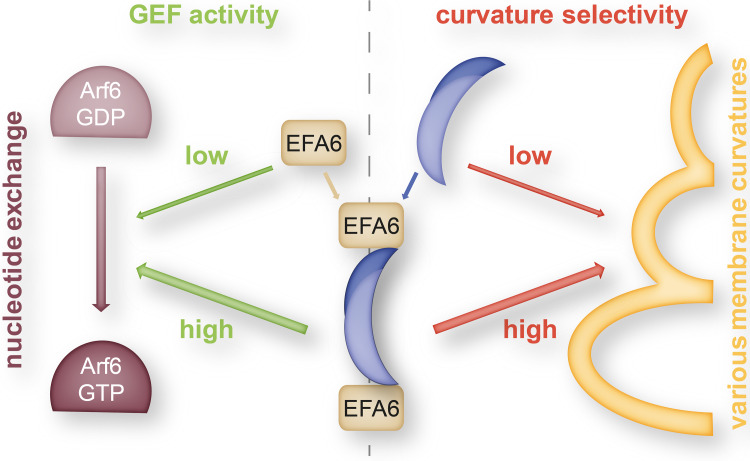
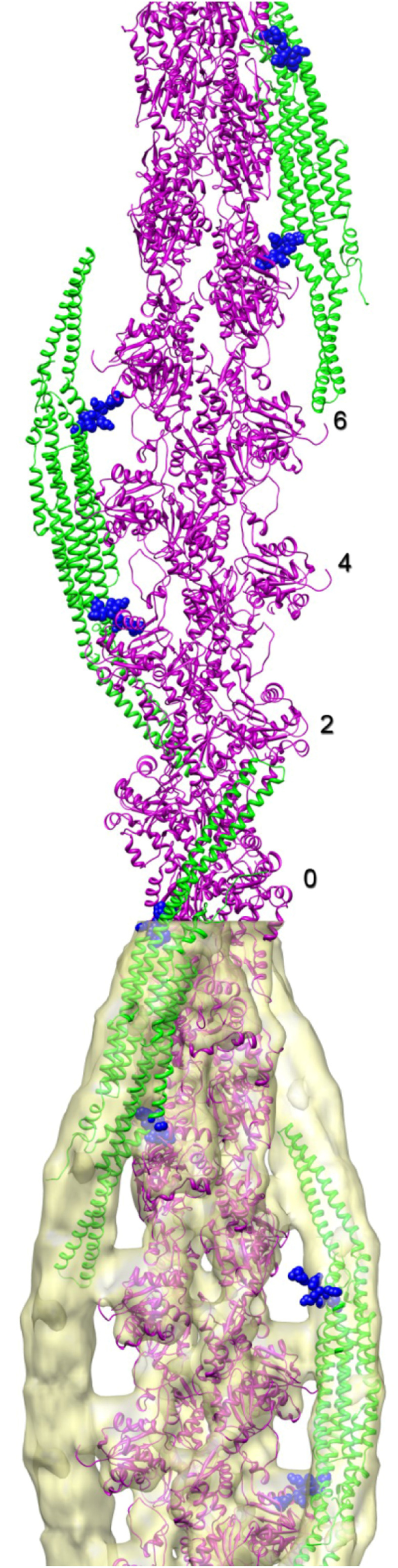
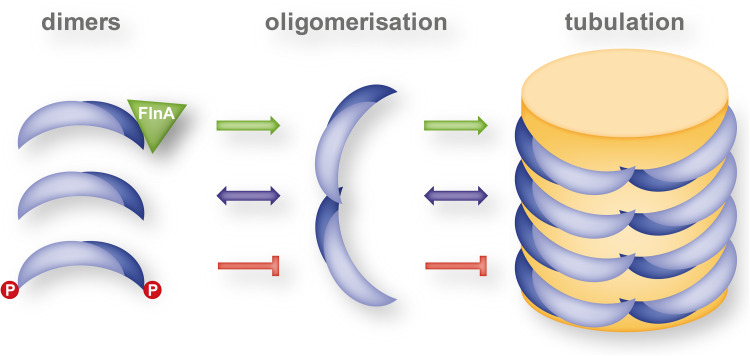
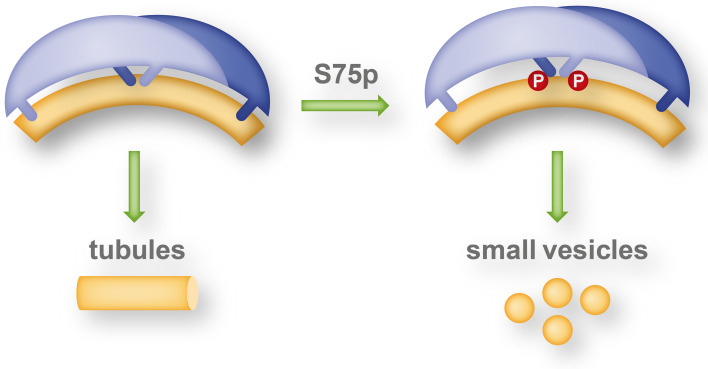
The BAR domain is the eponymous domain of the "BAR-domain protein superfamily", a large and diverse set of mostly multi-domain proteins that play eminent roles at the membrane cytoskeleton interface. BAR domain homodimers are the functional units that peripherally associate with lipid membranes and are involved in membrane sculpting activities. Differences in their intrinsic curvatures and lipid-binding properties account for a large variety in membrane modulating properties. Membrane activities of BAR domains are further modified and regulated by intramolecular or inter-subunit domains, by intermolecular protein interactions, and by posttranslational modifications. Rather than providing detailed cell biological information on single members of this superfamily, this review focuses on biochemical, biophysical, and structural aspects and on recent findings that paradigmatically promote our understanding of processes driven and modulated by BAR domains.
Keywords: F-BAR domain; I-BAR domain; Membrane curvature; Membrane remodelling; N-BAR domain; lipid binding.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Tsujita K, Suetsugu S, Sasaki N, Furutani M, Oikawa T, Takenawa T. Coordination between the actin cytoskeleton and membrane deformation by a novel membrane tubulation domain of PCH proteins is involved in endocytosis. J Cell Biol. 2006;172(2):269–279. doi: 10.1083/jcb.200508091. - DOI - PMC - PubMed
-
- Suetsugu S, Murayama K, Sakamoto A, Hanawa-Suetsugu K, Seto A, Oikawa T, Mishima C, Shirouzu M, Takenawa T, Yokoyama S. The RAC binding domain/IRSp53-MIM homology domain of IRSp53 induces RAC-dependent membrane deformation. J Biol Chem. 2006;281(46):35347–35358. doi: 10.1074/jbc.M606814200. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
