

Linking the community structure of arbuscular mycorrhizal fungi and plants: a story of interdependence?
- PMID: 28244977
- PMCID: PMC5437357
- DOI: 10.1038/ismej.2017.5
Linking the community structure of arbuscular mycorrhizal fungi and plants: a story of interdependence?
Abstract
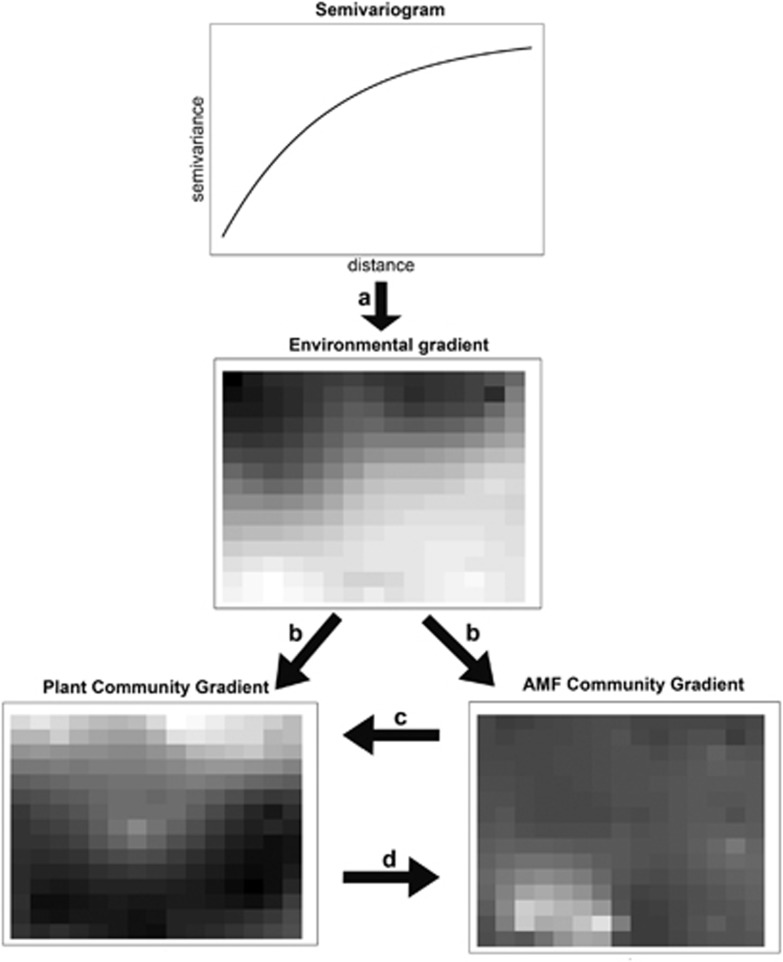
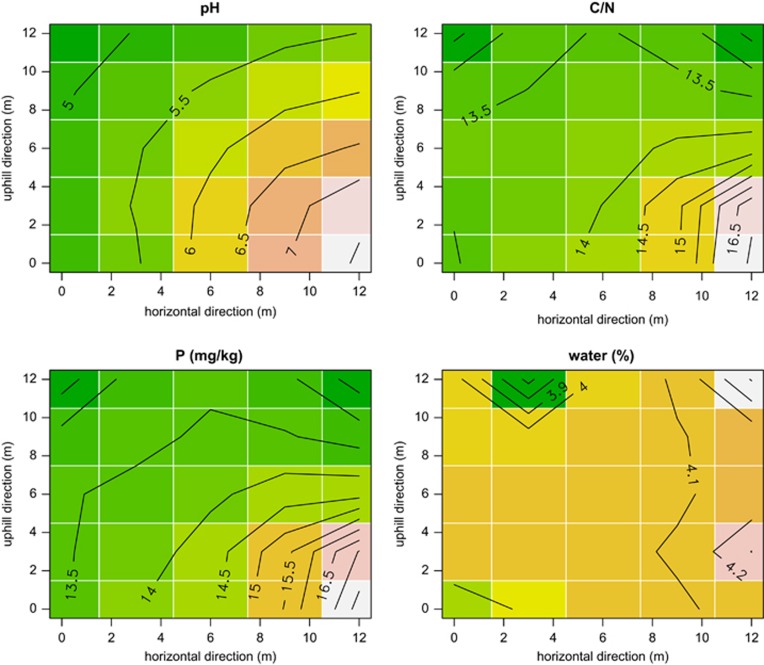
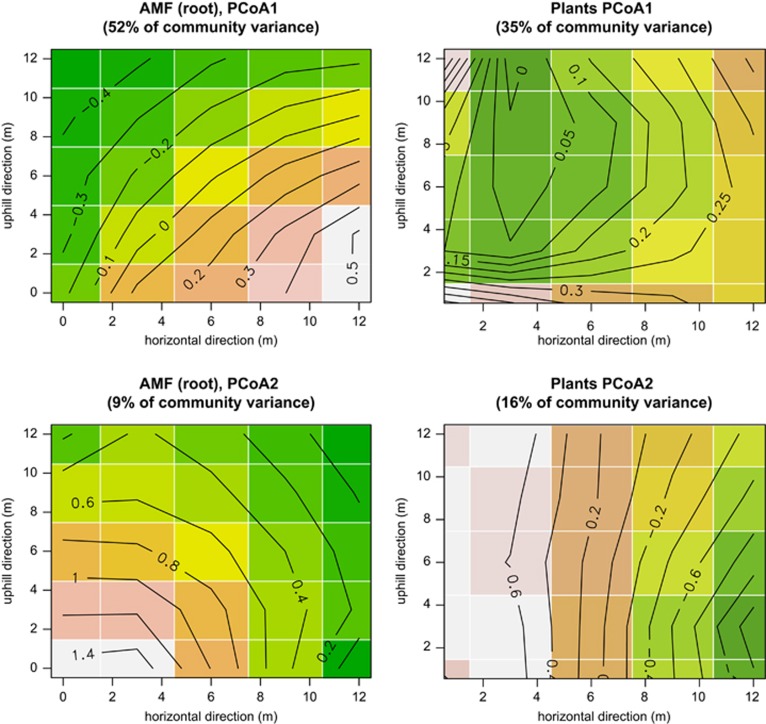
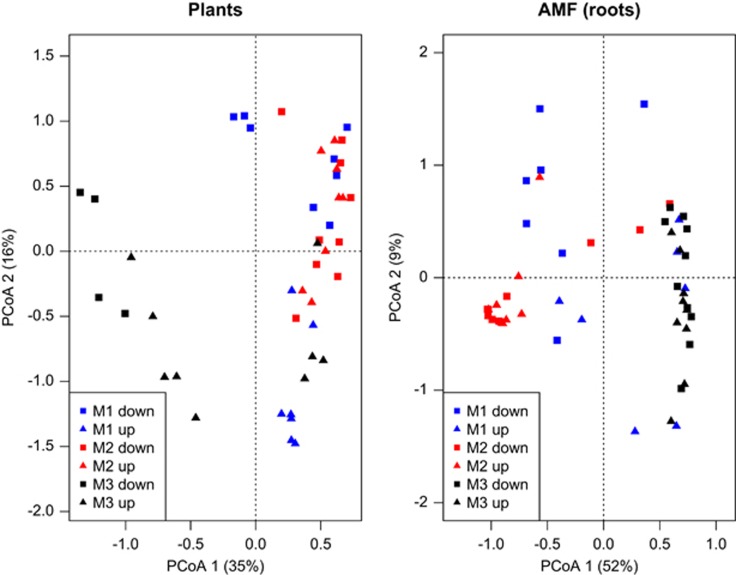
Arbuscular mycorrhizal fungi (AMF) are crucial to plants and vice versa, but little is known about the factors linking the community structure of the two groups. We investigated the association between AMF and the plant community structure in the nearest neighborhood of Festuca brevipila in a semiarid grassland with steep environmental gradients, using high-throughput sequencing of the Glomeromycotina (former Glomeromycota). We focused on the Passenger, Driver and Habitat hypotheses: (i) plant communities drive AMF (passenger); (ii) AMF communities drive the plants (driver); (iii) the environment shapes both communities causing covariation. The null hypothesis is that the two assemblages are independent and this study offers a spatially explicit novel test of it in the field at multiple, small scales. The AMF community consisted of 71 operational taxonomic units, the plant community of 47 species. Spatial distance and spatial variation in the environment were the main determinants of the AMF community. The structure of the plant community around the focal plant was a poor predictor of AMF communities, also in terms of phylogenetic community structure. Some evidence supports the passenger hypothesis, but the relative roles of the factors structuring the two groups clearly differed, leading to an apparent decoupling of the two assemblages at the relatively small scale of this study. Community phylogenetic structure in AMF suggests an important role of within-assemblage interactions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Alguacil MM, Torrecillas E, Garcia-Orenes F, Roldan A. (2014). Changes in the composition and diversity of AMF communities mediated by management practices in a Mediterranean soil are related with increases in soil biological activity. Soil Biol Biochem 76: 34–44.
-
- Anacker BL, Klironomos JN, Maherali H, Reinhart KO, Strauss SY. (2014). Phylogenetic conservatism in plant-soil feedback and its implications for plant abundance. Ecol Lett 17: 1613–1621. - PubMed
-
- Bever JD, Richardson SC, Lawrence BM, Holmes J, Watson M. (2009). Preferential allocation to beneficial symbiont with spatial structure maintains mycorrhizal mutualism. Ecol Lett 12: 13–21. - PubMed
-
- Camenzind T, Hempel S, Homeier J, Horn S, Velescu A, Wilcke W et al. (2014). Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest. Glob Chang Biol 20: 3646–3659. - PubMed
-
- Caravaca F, Ruess L. (2014). Arbuscular mycorrhizal fungi and their associated microbial community modulated by Collembola grazers in host plant free substrate. Soil Biol Biochem 69: 25–33.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
