

GTPase ROP2 binds and promotes activation of target of rapamycin, TOR, in response to auxin
- PMID: 28246118
- PMCID: PMC5376970
- DOI: 10.15252/embj.201694816
GTPase ROP2 binds and promotes activation of target of rapamycin, TOR, in response to auxin
Abstract
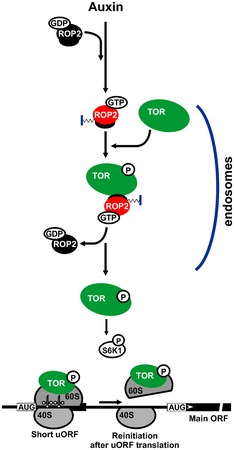
Target of rapamycin (TOR) promotes reinitiation at upstream ORFs (uORFs) in genes that play important roles in stem cell regulation and organogenesis in plants. Here, we report that the small GTPase ROP2, if activated by the phytohormone auxin, promotes activation of TOR, and thus translation reinitiation of uORF-containing mRNAs. Plants with high levels of active ROP2, including those expressing constitutively active ROP2 (CA-ROP2), contain high levels of active TOR ROP2 physically interacts with and, when GTP-bound, activates TOR in vitro TOR activation in response to auxin is abolished in ROP-deficient rop2 rop6 ROP4 RNAi plants. GFP-TOR can associate with endosome-like structures in ROP2-overexpressing plants, indicating that endosomes mediate ROP2 effects on TOR activation. CA-ROP2 is efficient in loading uORF-containing mRNAs onto polysomes and stimulates translation in protoplasts, and both processes are sensitive to TOR inhibitor AZD-8055. TOR inactivation abolishes ROP2 regulation of translation reinitiation, but not its effects on cytoskeleton or intracellular trafficking. These findings imply a mode of translation control whereby, as an upstream effector of TOR, ROP2 coordinates TOR function in translation reinitiation pathways in response to auxin.
Keywords: S6K1; endosomes; phosphorylation; phytohormone auxin; signal transduction.
© 2017 The Authors.
Figures

Schematic representation of Arabidopsis TOR (S2424 phosphorylation site indicated) and ROP functional domains (G domains, the positions of Q64 and D121, and C‐terminal basic K/R‐CaaL motifs are indicated).
Phylogenetic tree of 11 Rop family member proteins. ROPs 2, 4, and 6 are classified in a subgroup (red).
ROP2, ROP4, and ROP6 identified as putative TOR interactors by the yeast two‐hybrid (Y2H) system. BD‐ROPs 1–6 were assayed for interaction with AD‐TOR. Equal OD600 units and 1/10 and 1/100 dilutions were spotted from left to right.
GST pull‐down assay: ROP2‐, Rheb‐, Sar1b‐tagged GST, and GST alone were assayed for interaction with recombinant TOR as indicated on the left panel. GST‐fusion protein bound (B) and unbound (U) fractions were stained by Coomassie blue.
Immunoprecipitation (IP) experiments with anti‐GFP‐Trap magnetic beads on crude extracts of GFP‐TOR and GFP transgenic plants; for Western blots, 10% of the input and 100% of IP fractions were analyzed with anti‐GFP, anti‐TOR, and anti‐ROP antibodies (Abs).
Endogenous TOR was immunoprecipitated from Arabidopsis extract with anti‐TOR Abs (IP) and assayed for association with ROPs by immunoblotting. 10% of the input and 100% of IP or normal rabbit serum (NRS) were analyzed by anti‐ROP antibodies.
Y2H: TOR and its N‐terminal domain (NTOR) interact with ROP2 and dominant‐negative ROP2 (DN‐ROP2). AD‐TOR, ‐NTOR, and ‐CTOR were assayed for interaction with BD‐ROP2 or ‐ROP2 mutants ‐CA‐ROP2 and ‐DN‐ROP2 as indicated.
Left panel: GST pull‐down assay: ROP2‐, DN‐ROP2‐, CA‐ROP2‐tagged GST, and GST alone were assayed for interaction with recombinant TOR. Fractions were stained by Coomassie blue. Right panel: Quantification of TOR binding to GST‐fusion proteins. The value for TOR binding to GST‐ROP2 was set as 100%. Statistical analysis is based on one‐way ANOVA test. Data are presented as mean ± SEM (P < 0.05, n = 3).

ROPs 1–6 transcription profiles were taken from the Genevestigator database (
http://www.genevestigator.ethz.ch ).Alignment of the C‐terminal tail patterns of ROPs 1–6 from Arabidopsis and human RAC1. Two motifs are indicated: basic lysine residues (motif I), and a CxxL (x = aliphatic amino acid) geranylgeranylation motif (motif II). Two deletion variants used are indicated by solid lines. Alignment done in agreement with BLOSUM 62 and Jonson amino acid substitution matrices (similar residues are printed in reverse type).
GST pull‐down assay: ROP2‐, ROP2ΔII‐, ROP2Δ(I+II)‐tagged GST, and GST alone were assayed for interaction with recombinant TOR. Fractions were stained by Coomassie blue. Right panel: Quantification of TOR binding to GST‐fusion proteins. Results shown represent the means obtained in three independent experiments, and error bars indicate SD. The value for TOR binding to GST‐ROP2 was set as 1.

WT seedlings were treated with either NAA or AZD‐8055, or TOR was inactivated by AZD‐8055 in seedlings treated with NAA for 8 h. TOR levels and its phosphorylation status were analyzed by immunoblot with anti‐AtTOR Abs (anti‐TOR) and anti‐(mTOR‐S2448‐P) Abs, respectively. Loading control was stained with Coomassie blue. Right: Quantification of ratio between TOR‐P and TOR. The value for TOR‐P/TOR in WT was set as 1.
Rosettes representative of WT, yuc1D, and cuf1D plants.
Analysis of active TOR and ROPs‐GTP levels in WT, yuc1D, and cuf1D 7‐dag seedlings. Input: Total ROPs, TOR total, and its phosphorylation levels were analyzed by Western blot. GST‐Ric1 and loading control were stained with Coomassie blue. Bottom left: Quantification of ratio between TOR‐P and TOR (n = 3). The value for TOR‐P/TOR in WT was set as 1. Right panels: Analysis of active ROPs by GST‐Ric1 IP in seedling extracts. GST‐Ric1‐bound ROPs were detected using anti‐ROP antibodies. Quantification of ratio between ROPs‐GTP and GST‐Ric1. The value for ROPs‐GTP/GST‐Ric1 in WT was set as 1.

Rosettes representative of WT, cuf1D, and CA‐ROP2 plants.
GST‐Ric1 (or GST) pull‐down IP assays targeting active GTP‐bound ROPs in WT, cuf1D, and CA‐ROP2 seedlings. Active ROPs and total ROPs were detected with an anti‐ROP Abs. GST‐Ric1 and a loading control (LC) were stained with Coomassie blue.
The level of endogenous mRNAs, including actin (ACT), glycerol‐3‐phosphate dehydrogenase C2 (GAPC2), expressed protein (EXP), and others indicated below the bar graphs in WT and cuf1D, was examined by qRT–PCR. The RNA value in WT extracts was set as 100%.
TOR and S6K1 levels and their phosphorylation status in either cuf1D and WT, or CA‐ROP2 and WT were analyzed by immunoblot with anti‐AtTOR Abs (anti‐TOR), anti‐(mTOR‐S2448‐P) and anti‐mS6K1, and anti‐(mS6K1‐T389‐P) Abs, respectively. The density of bands on Western blots was quantified, and WT values were set as 100%.
Left panel: Analysis of active ROPs by GST‐Ric1 IP in WT and CA‐ROP2 extracts from 7‐dag seedlings grown with or without 1 μM AZD‐8055. Total ROPs and GST‐Ric1 bound ROPs were detected using anti‐ROP antibodies. GST‐Ric1 and LC were stained with Coomassie blue. Input: Total ROPs, TOR total, and its phosphorylation levels were analyzed by Western blot. Right panel: Quantification of ratio between ROPs‐GTP and GST‐Ric1. The value for ROPs‐GTP in WT and CA‐ROP2 was set as 1.
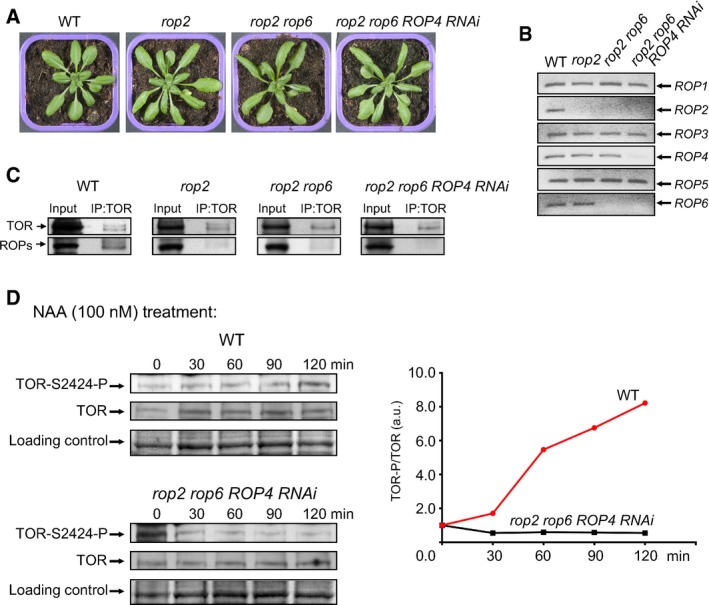
Rosettes representative of WT, rop2, rop2 rop6, and rop2 rop6 ROP4 RNAi plants.
The level of endogenous ROP mRNAs in different ROP‐deficient plants was examined by a semiquantitative RT–PCR.
Endogenous TOR was immunoprecipitated from WT and ROP‐deficient Arabidopsis extracts by immunoblotting with anti‐TOR Abs (IP) and assayed for association with ROPs. 10% of the input and 100% of IP were analyzed by anti‐ROP antibodies.
Time‐course of TOR and TOR‐P accumulation in extracts from 7‐dag seedlings before (0 min) and after transfer to medium with NAA analyzed by immunoblot with anti‐TOR and anti‐(mTOR‐S2448‐P) Abs. The value of TOR‐P/TOR at 0 min for each line was set as 1.

Rosettes representative of WT, GFP‐TOR, CA‐ROP2, and GFP‐TOR/CA‐ROP2 plants.
The level of endogenous mRNAs including endogenous TOR (TOR end) and both GFP‐TOR and TOR end (TOR mix) and others indicated below the bar graphs in GFP‐TOR and GFP‐TOR/CA‐ROP2 was examined by qRT–PCR. The RNA value in GFP‐TOR extracts was set as 100%. Values, expressed in arbitrary units, are averages of three technical replicates, and error bars indicate SD.
Total and active ROP and TOR levels, and TOR phosphorylation status in either GFP‐TOR or GFP‐TOR/CA‐ROP2 were analyzed as described in Fig 2B and D, respectively.
In vitro phosphorylation kinetics of recombinant S6K1 (S6K1) using GFP‐TOR immunoprecipitated from GFP‐TOR or GFP‐TOR/CA‐ROP2. S6K1 total and phosphorylation levels were followed by Western blot using anti‐S6K1‐T389‐P or anti‐S6K1 Abs. Total S6K1 was stained by Coomassie blue.
Representative images of pavement cell (PC) morphology in the second true leaf of 21‐day‐old WT plants and the GFP‐TOR, CA‐ROP2, and GFP‐TOR/CA‐ROP2 mutant lines. Scale bars are 30 μm. Bottom panel: Quantitative analysis of cell circularity (left) and cell sizes (right) is shown as box and whisker plots with median, lower Q1 and upper Q3 quartiles, error bars indicating minimum to maximum range, and presented using unpaired t‐test. *P < 0.05; **P < 0.001; ns, non‐significant (n = 46).

- A
Nicotiana benthamiana epidermal cells transiently expressing GFP‐TOR (left panel) and co‐transformed (from left to right) with myc‐ROP2, or myc‐CA‐ROP2, or myc‐DN‐ROP2. Quantitative analysis of GFP‐TOR aggregate number (box and whiskers plot, left; P < 0.05, n = 6) and sizes (scatter plot, right; P < 0.0001, n = 6). The box and whisker plot is with median, lower Q1 and upper Q3 quartiles, error bars indicating minimum to maximum range. Statistical analysis is based on one‐way ANOVA test.
- B
Immunoblot analysis with anti‐myc or anti‐GFP of transiently co‐expressed GFP‐TOR without or with myc‐ROP2, myc‐CA‐ROP2, or myc‐DN‐ROP2 in N. benthamiana cells.
- C–F
Fluorescence micrographs showing N. benthamiana cells transiently expressing: (C) GFP‐TOR; (D) GFP‐TOR (upper left), RFP‐ROP2 (upper center), GFP‐TOR (middle left), RFP‐CA‐ROP2 (middle center), GFP‐TOR (bottom left), and RFP‐DN‐ROP2 (bottom center); (E) RFP‐ROP2 (left), RFP‐ROP2∆II (CAFL) (center), and RFP‐ROP2Δ(I+II) (right); (F) GFP‐TOR (upper left), RFP‐ROP2 (upper center), GFP‐TOR (middle left), RFP‐ROP2ΔII (middle center), GFP‐TOR (bottom left), and RFP‐ROP2Δ(I+II) (bottom center). Merged images are shown on the right.
- G
Imaging fluorescence assays showing root cells of Arabidopsis 7‐dag GFP‐TOR and GFP‐TOR/CA‐ROP2 cells.
- H
Intracellular distribution of TOR and active TOR. Western blot analysis of various fractions following microsome isolation from GFP‐TOR and GFP‐TOR/CA‐ROP2. The total homogenate (total), nuclear fraction pellet (P10), pellet (P30), pellet (P100), supernatant (S100) were analyzed by Western blot with corresponding antibodies.

Fluorescence micrographs showing Nicotiana benthamiana cells transiently expressing plasma membrane (PM) marker GFP‐BD‐CVIL (left panels), RFP‐ROP2 (upper center), RFP‐ROP2ΔII (middle center), and RFP‐ROP2Δ(I+II) (bottom center). Merged images are shown on the right. PM marker consists of green fluorescent protein (GFP) fused to C‐terminal polybasic domain (BD) and isoprenylation motif (CVIL).
Imaging fluorescence assays showing root cells of Arabidopsis 7‐dag GFP and GFP‐TOR seedlings before and after treatment with 100 nM NAA.
Intracellular distribution of TOR and active TOR in WT seedlings before (left panel) and after treatment by 100 nM NAA for 8 h (right panel). Western blot analysis of various fractions: the total homogenate (total), nuclear fraction pellet (P10), 30,000 g pellet (P30), 100,000 g pellet (P100), 100,000 g supernatant (S100).
Nicotiana benthamiana cells transiently co‐expressing GFP‐TOR, RFP‐Golgi, or both with either myc‐DN‐Sar1b or myc‐DN‐ROP2.
Immunoblot analysis with anti‐myc Abs of cells.

- A–G
Imaging fluorescence assays showing Nicotiana benthamiana cells transiently co‐expressing FLAG‐CA‐ROP2 with GFP‐TOR (left panels), and intracellular markers (central panels) that specifically label early endosomes (A, RFP‐RabC1), endosomes (B, RFP‐RabE1d), autophagosomes (C, RFP‐ATG8a), peroxisomes (mCherry‐peroxi), mitochondria (E, mCherry‐mito), late endosomes (F, RFP‐ARA7), and Golgi (G, GmMan1‐tdTomato); merged images (right). Scale bars are 5 μm.

Co‐localization analysis of GFP‐TOR and RFP‐RabC1 in Nicotiana benthamiana epidermal cells expressing FLAG‐ROP2 (upper panels), FLAG‐DN‐ROP2 (central panels), and FLAG‐CA‐ROP2 (bottom panels).
35S:RFP‐RabC1 Arabidopsis line transiently expressing GFP‐TOR and FLAG‐CA‐ROP2. GFP‐TOR (left), RFP‐Rab1C (center), merged image (right).
35S:GFP‐TOR Arabidopsis line transiently expressing RFP‐RabC1 and FLAG‐CA‐ROP2. GFP‐TOR (left), RFP‐RabC1 (center), merged image (right).
Microscopy images of cells stained with FM4‐64 treated with brefeldin A (BFA) in the root elongation zone of GFP‐TOR 7‐dag seedlings. GFP‐TOR and FM4‐64 were detected in the core of the BFA compartment.
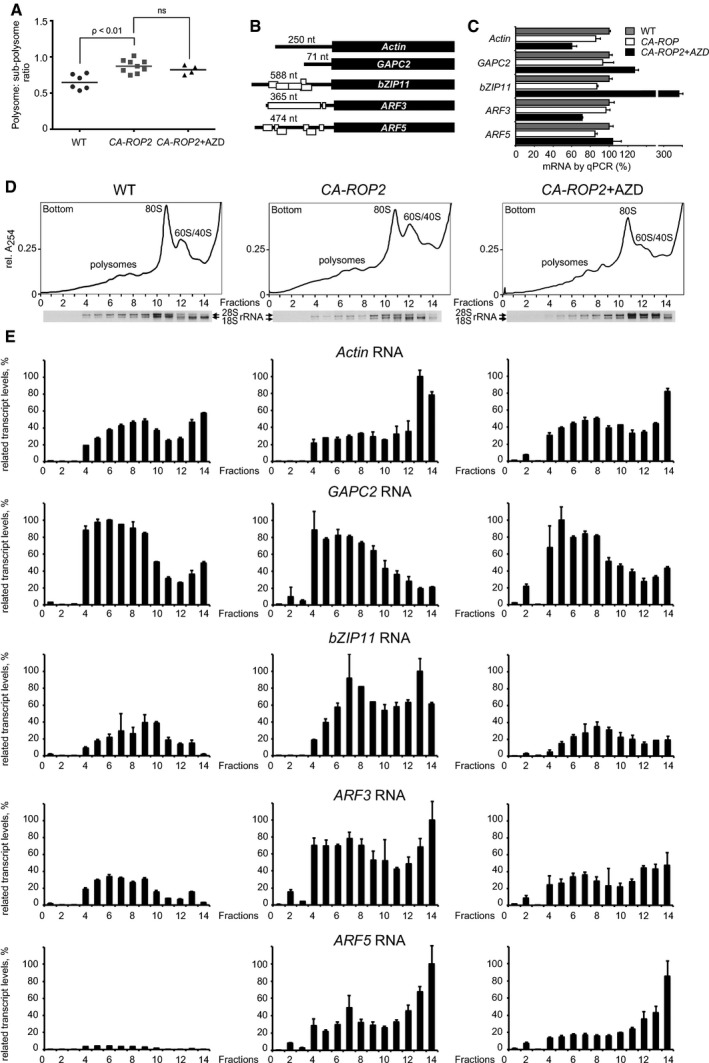
Statistical analyses of ratio between polysomal and non‐polysomal fractions obtained by sucrose gradient fractionation of extracts isolated from WT seedlings and CA‐ROP2 seedlings grown without or with 0.5 μM AZD‐8055.
uORF (open rectangles) configuration within selected mRNAs.
qRT–PCR of each mRNA in total extracts. The RNA value in WT extracts was set as 100%.
Ribosome sedimentation profiling from extracts prepared from WT (left panel) and CA‐ROP2 7‐dag seedlings treated (right panel) or not (central panel) with 0.5 μM AZD‐8055. Positions of ribosomal subunits (60S/40S), monosomes (80S), and polysomes are indicated. 18S and 28S rRNA distribution was monitored by agarose gel electrophoresis.
mRNA association with polyribosomes, 80S, and 60S/40S ribosomal subunits was monitored by qRT–PCR in sucrose gradient fractions and presented as graph bars. The highest value of each selected polysome‐bound mRNA among WT, CA‐ROP2, and CA‐ROP2 + AZD was set as 100%.

qRT–PCR of each mRNA in total extracts. The RNA value in WT extracts was set as 100%.
The level of heavy polysomes is reduced in WT Arabidopsis treated by AZD‐8055 (AZD). Extracts prepared from 7‐dag seedlings growing without (WT) and with 0.5 μM AZD‐8055 on agar plates (WT + AZD) were subjected to velocity sedimentation through sucrose density gradients. Gradients were fractionated while scanning at 254 nm, and the resulting absorbance profiles are shown (WT and WT + AZD). Positions of ribosomal subunits (RS), monosomes (80S), and polysomes are indicated. rRNA distribution was monitored by agarose gel electrophoresis.
AZD‐8055 treatment down‐regulates, albeit not significantly, the abundance of bZIP11 and the already low polysomal levels of ARF5 mRNA in WT Arabidopsis. Distribution of mRNAs (actin, GAPC2, bZIP11, ARF3, and ARF5) in fractions were analyzed by qRT–PCR. Note that the highest value of each polysome‐bound mRNA from WT, CA‐ROP2, CA‐ROP2 + AZD (Fig 7) was used as 100% for both WT and WT + AZD.
TOR and TOR phosphorylation status were analyzed in polysomes prepared from WT, CA‐ROP2 seedlings and CA‐ROP2 line treated by AZD‐8055. Three samples from polysomes and two from 80S and ribosomal subunits were taken to monitor TOR by immunoblotting with anti‐TOR (lower panels) and phospho‐TOR with anti‐(mTOR‐S2448‐P) (central panels). Data shown are representative of two independent blots.

Phosphorylation of TOR at S2424 is augmented in CA‐ROP2‐overexpressing versus WT mesophyll protoplasts and diminished in the presence of 0.5 μM AZD‐8055. TOR and its phosphorylation levels were assayed by immunoblotting. The Western blot density bands were quantified and CA‐ROP2 values were set as 100%.
GUS‐containing reporters with either short or uORF‐containing (ARF3 and ARF5) 5′‐UTRs were used for mesophyll protoplasts transformation.
WT and CA‐ROP2 seedlings growing without (CA‐ROP2) or with 1 μM AZD‐8055 (CA‐ROP2 + AZD) were used to prepare mesophyll protoplasts. Both GFP fluorescence and β‐glucuronidase functional activity were analyzed in the same 96‐well microtiter plate. Functional levels of GUS expressed from pshort‐GUS normalized to corresponding GFP levels were set at 100%. GUS‐containing mRNA levels and integrity were analyzed by semiquantitative RT–PCR (sqRT–PCR); GFP levels were also analyzed by immunoblotting. LC, loading control.
pmonoGFP and either pARF3‐GUS or pARF3Δ(AUG1+2)‐GUS were used to transform WT or CA‐ROP2‐derived protoplasts. The GUS/GFP ratio found in WT protoplasts with a uORF‐less ARF3 leader was set as 100%.
WT mesophyll protoplasts were transfected in addition to pmonoGFP/pshort‐GUS or pARF5‐GUS with the vector expressing either myc‐tagged ROP2, CA‐ROP2, or DN‐ROP2 under the 35S promoter. Both GFP fluorescence and β‐glucuronidase functional activity were analyzed in the same 96‐well microtiter plate. GUS/GFP ratio related to the short or ARF5 5′‐UTRs was taken as 100%. GUS mRNA levels and integrity were analyzed by sqRT–PCR. ROP2 variants analyzed by immunoblotting using anti‐myc Abs (samples were run on the same gel and the solid line indicates removal of extraneous lanes).
Dose‐dependent effect of AZD‐8055 on root growth of WT and CA‐ROP2 seedlings at 24, 48, and 72 h.
Dose‐dependent effect of AZD‐8055 on root hair elongation of WT and CA‐ROP2 seedlings.
Similar articles
-
TOR and S6K1 promote translation reinitiation of uORF-containing mRNAs via phosphorylation of eIF3h.EMBO J. 2013 Apr 17;32(8):1087-102. doi: 10.1038/emboj.2013.61. Epub 2013 Mar 22. EMBO J. 2013. PMID: 23524850 Free PMC article.
-
Diverse nitrogen signals activate convergent ROP2-TOR signaling in Arabidopsis.Dev Cell. 2021 May 3;56(9):1283-1295.e5. doi: 10.1016/j.devcel.2021.03.022. Epub 2021 Apr 7. Dev Cell. 2021. PMID: 33831352
-
COP1 integrates light signals to ROP2 for cell cycle activation.Plant Signal Behav. 2017 Sep 2;12(9):e1363946. doi: 10.1080/15592324.2017.1363946. Epub 2017 Aug 14. Plant Signal Behav. 2017. PMID: 28805486 Free PMC article.
-
Auxin Signaling in Regulation of Plant Translation Reinitiation.Front Plant Sci. 2017 Jun 14;8:1014. doi: 10.3389/fpls.2017.01014. eCollection 2017. Front Plant Sci. 2017. PMID: 28659957 Free PMC article. Review.
-
From perception to attenuation: auxin signalling and responses.Curr Opin Plant Biol. 2013 Oct;16(5):561-8. doi: 10.1016/j.pbi.2013.08.003. Epub 2013 Sep 1. Curr Opin Plant Biol. 2013. PMID: 24004572 Review.
Cited by
-
Transcriptomic and splicing changes underlying tomato responses to combined water and nutrient stress.Front Plant Sci. 2022 Nov 25;13:974048. doi: 10.3389/fpls.2022.974048. eCollection 2022. Front Plant Sci. 2022. PMID: 36507383 Free PMC article.
-
Metabolic regulation of quiescence in plants.Plant J. 2023 Jun;114(5):1132-1148. doi: 10.1111/tpj.16216. Epub 2023 Apr 13. Plant J. 2023. PMID: 36994639 Free PMC article.
-
Converging Light, Energy and Hormonal Signaling Control Meristem Activity, Leaf Initiation, and Growth.Plant Physiol. 2018 Feb;176(2):1365-1381. doi: 10.1104/pp.17.01730. Epub 2017 Dec 28. Plant Physiol. 2018. PMID: 29284741 Free PMC article.
-
Do plants drive translation reinitiation to dodge nonsense-mediated decay?J Exp Bot. 2023 Jan 1;74(1):7-11. doi: 10.1093/jxb/erac444. J Exp Bot. 2023. PMID: 36563104 Free PMC article.
-
Translational fidelity and growth of Arabidopsis require stress-sensitive diphthamide biosynthesis.Nat Commun. 2022 Jul 11;13(1):4009. doi: 10.1038/s41467-022-31712-7. Nat Commun. 2022. PMID: 35817801 Free PMC article.
References
-
- Andreeva AV, Zheng H, Saint‐Jore CM, Kutuzov MA, Evans DE, Hawes CR (2000) Organization of transport from endoplasmic reticulum to Golgi in higher plants. Biochem Soc Trans 28: 505–512 - PubMed
-
- Beltrán‐Peña E, Aguilar R, Ortíz‐López A, Dinkova TD, De Jiménez ES (2002) Auxin stimulates S6 ribosomal protein phosphorylation in maize thereby affecting protein synthesis regulation. Physiol Plant 115: 291–297 - PubMed
-
- Berken A, Wittinghofer A (2008) Structure and function of Rho‐type molecular switches in plants. Plant Physiol Biochem 46: 380–393 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
