

Membrane Currents, Gene Expression, and Circadian Clocks
- PMID: 28246182
- PMCID: PMC5411696
- DOI: 10.1101/cshperspect.a027714
Membrane Currents, Gene Expression, and Circadian Clocks
Abstract
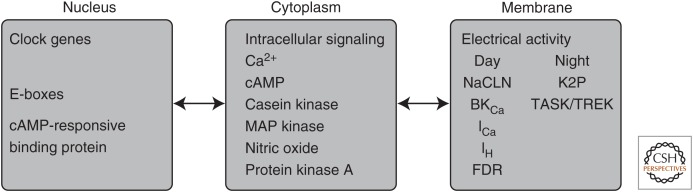
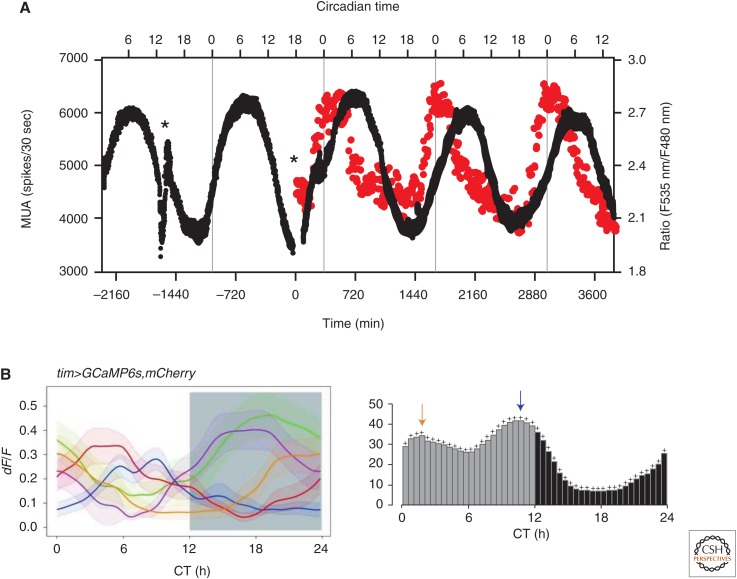
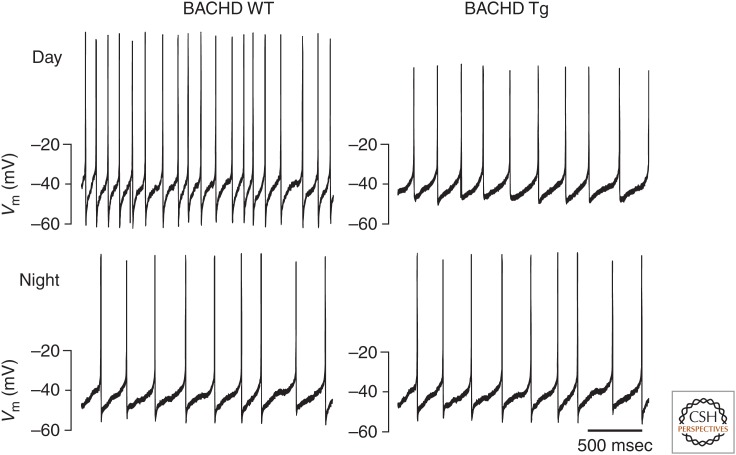
Neuronal circadian oscillators in the mammalian and Drosophila brain express a circadian clock comprised of interlocking gene transcription feedback loops. The genetic clock regulates the membrane electrical activity by poorly understood signaling pathways to generate a circadian pattern of action potential firing. During the day, Na+ channels contribute an excitatory drive for the spontaneous activity of circadian clock neurons. Multiple types of K+ channels regulate the action potential firing pattern and the nightly reduction in neuronal activity. The membrane electrical activity possibly signaling by changes in intracellular Ca2+ and cyclic adenosine monophosphate (cAMP) regulates the activity of the gene clock. A decline in the signaling pathways that link the gene clock and neural activity during aging and disease may weaken the circadian output and generate significant impacts on human health.
Copyright © 2017 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Akiyama M, Minami Y, Nakajima T, Moriya T, Shibata S. 2001. Calcium and pituitary adenylate cyclase-activating polypeptide induced expression of circadian clock gene mPer1 in the mouse cerebellar granule cell culture. J Neurochem 78: 499–508. - PubMed
-
- Albrecht U, Sun ZS, Eichele G, Lee CC. 1997. A differential response of two putative mammalian circadian regulators, mper1 and mper2, to light. Cell 91: 1055–1064. - PubMed
-
- Albus H, Bonnefont X, Chaves I, Yasui A, Doczy J, van der Horst GT, Meijer JH. 2002. Cryptochrome-deficient mice lack circadian electrical activity in the suprachiasmatic nuclei. Curr Biol 12: 1130–1133. - PubMed
-
- Asai M, Yoshinobu Y, Kaneko S, Mori A, Nikaido T, Moriya T, Akiyama M, Shibata S. 2001. Circadian profile of Per gene mRNA expression in the suprachiasmatic nucleus, paraventricular nucleus, and pineal body of aged rats. J Neurosci Res 66: 1133–1139. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous