

Metabolic origins of spatial organization in the tumor microenvironment
- PMID: 28246332
- PMCID: PMC5358370
- DOI: 10.1073/pnas.1700600114
Metabolic origins of spatial organization in the tumor microenvironment
Abstract
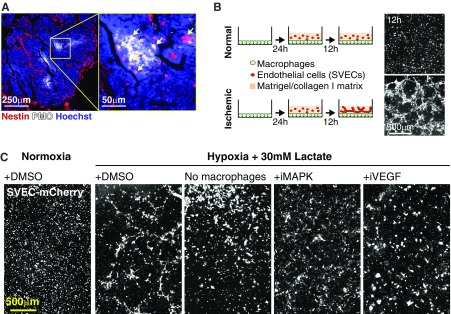
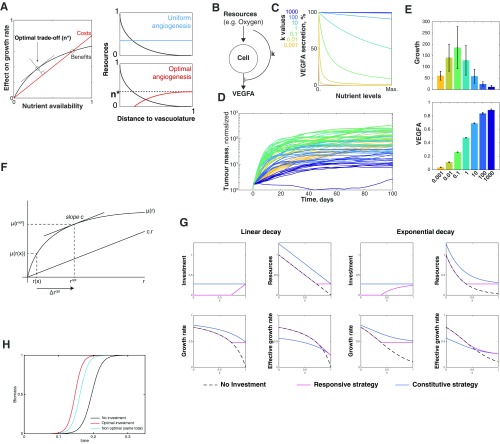
The genetic and phenotypic diversity of cells within tumors is a major obstacle for cancer treatment. Because of the stochastic nature of genetic alterations, this intratumoral heterogeneity is often viewed as chaotic. Here we show that the altered metabolism of cancer cells creates predictable gradients of extracellular metabolites that orchestrate the phenotypic diversity of cells in the tumor microenvironment. Combining experiments and mathematical modeling, we show that metabolites consumed and secreted within the tumor microenvironment induce tumor-associated macrophages (TAMs) to differentiate into distinct subpopulations according to local levels of ischemia and their position relative to the vasculature. TAMs integrate levels of hypoxia and lactate into progressive activation of MAPK signaling that induce predictable spatial patterns of gene expression, such as stripes of macrophages expressing arginase 1 (ARG1) and mannose receptor, C type 1 (MRC1). These phenotypic changes are functionally relevant as ischemic macrophages triggered tube-like morphogenesis in neighboring endothelial cells that could restore blood perfusion in nutrient-deprived regions where angiogenic resources are most needed. We propose that gradients of extracellular metabolites act as tumor morphogens that impose order within the microenvironment, much like signaling molecules convey positional information to organize embryonic tissues. Unearthing embryology-like processes in tumors may allow us to control organ-like tumor features such as tissue repair and revascularization and treat intratumoral heterogeneity.
Keywords: cancer metabolism; morphogens; positional information; tumor microenvironment; tumor-associated macrophages.
Conflict of interest statement
Conflict of interest statement: C.B.T. is a founder of Agios Pharmaceuticals and a member of its scientific advisory board. He also serves on the board of directors of Merck and Charles River Laboratories.
Figures
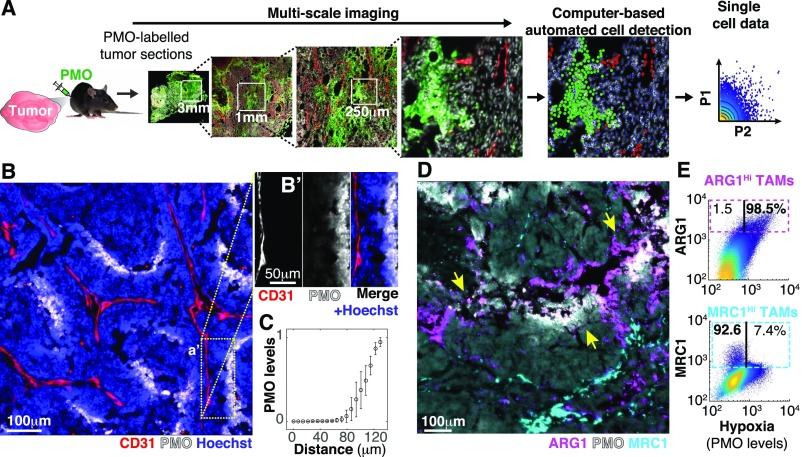
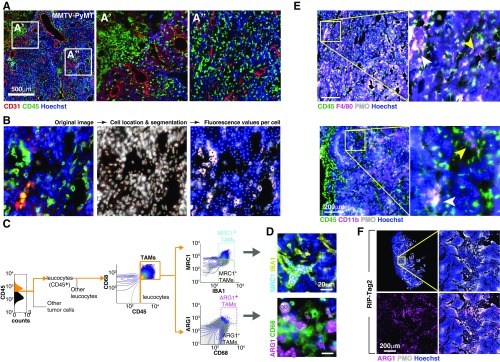
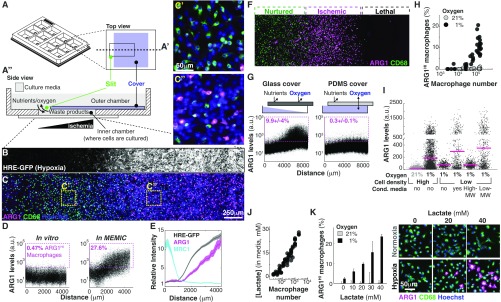
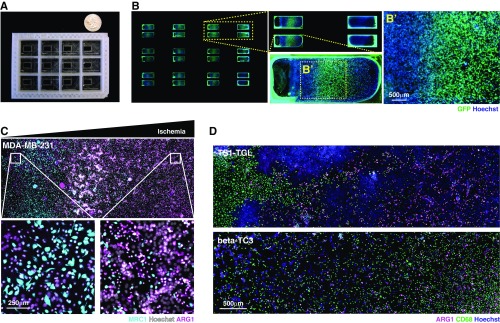
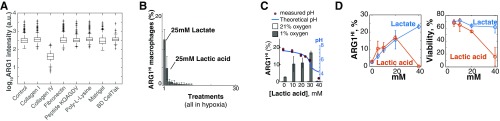
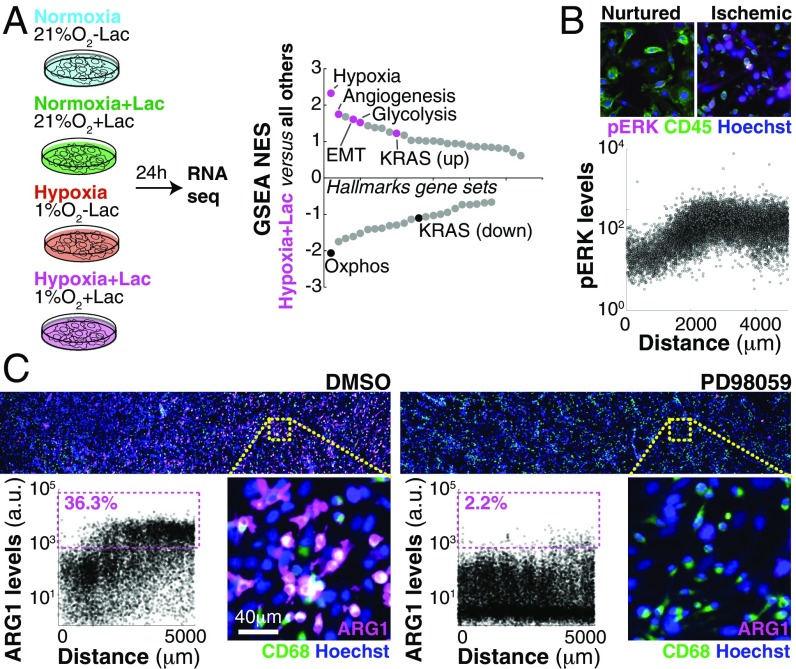
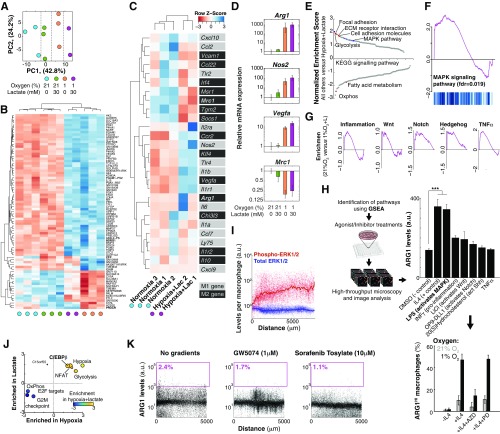
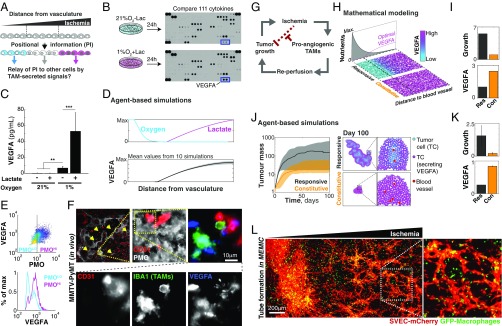
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
