

Translational Control of Germ Cell Decisions
- PMID: 28247049
- PMCID: PMC5985952
- DOI: 10.1007/978-3-319-44820-6_6
Translational Control of Germ Cell Decisions
Abstract
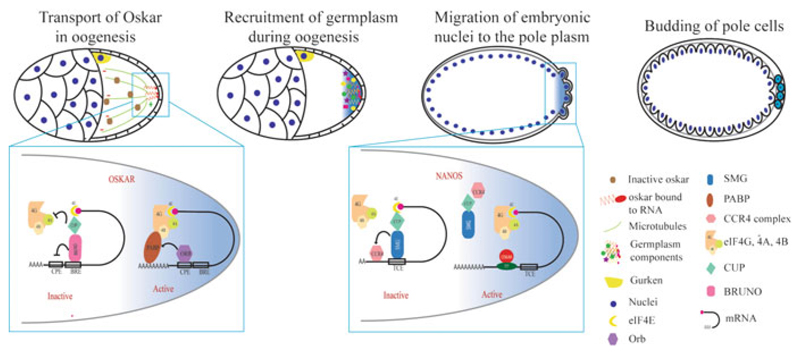
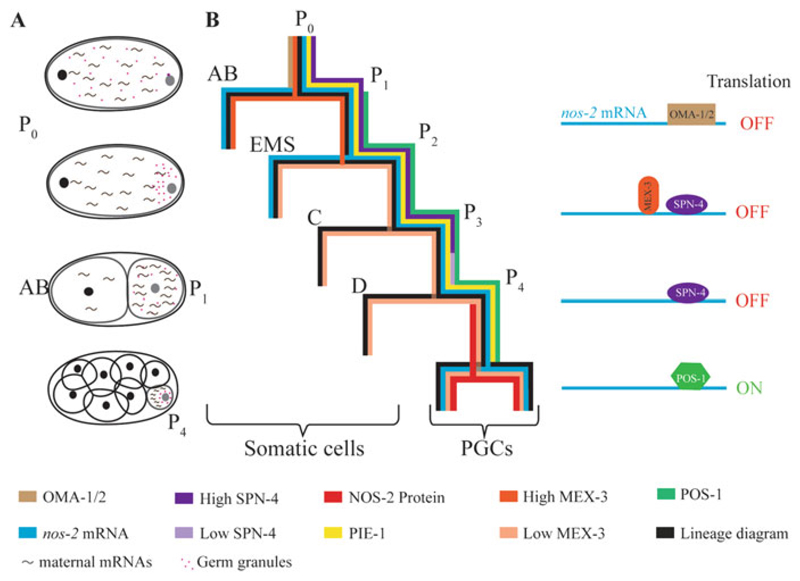
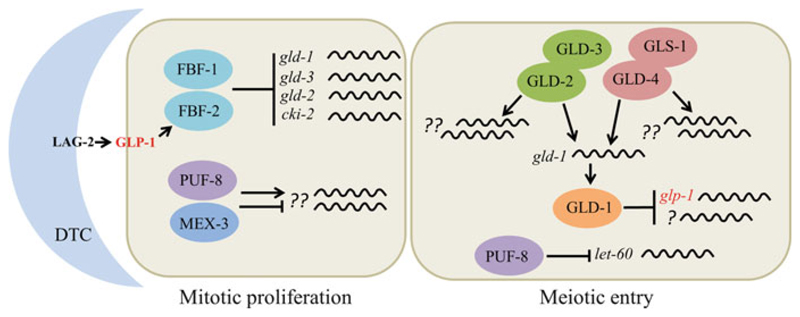
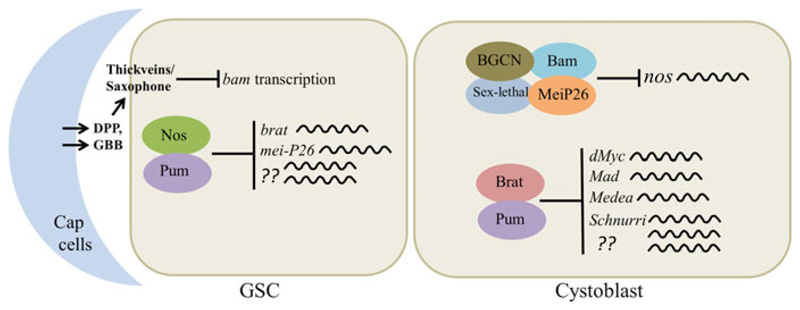
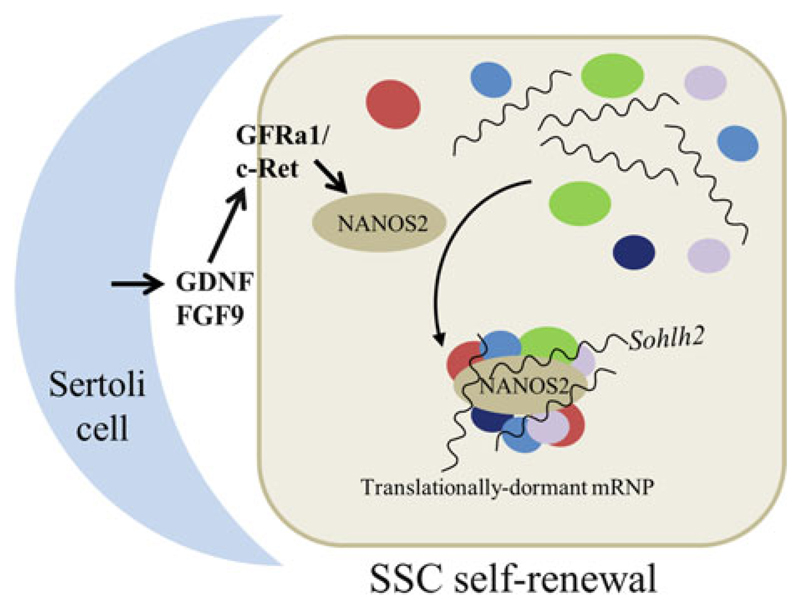
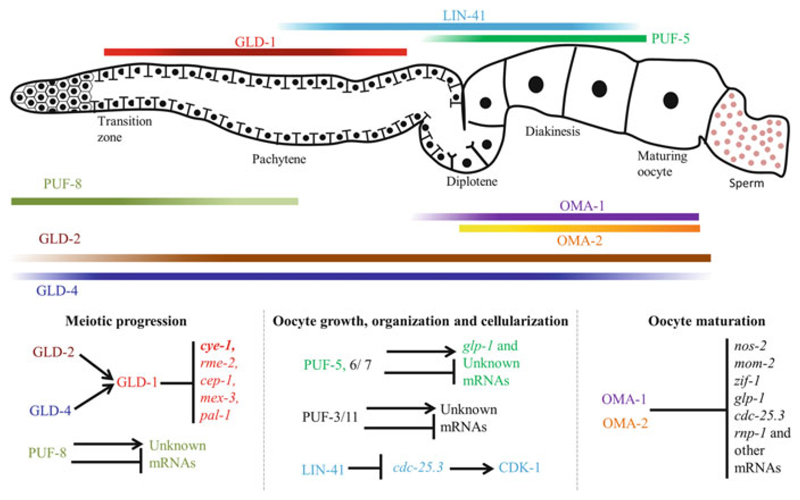
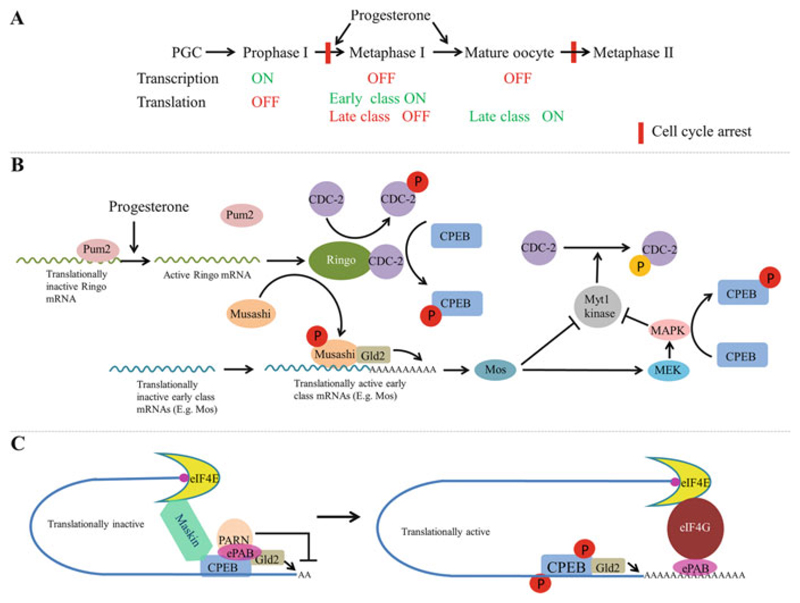
Germline poses unique challenges to gene expression control at the transcriptional level. While the embryonic germline maintains a global hold on new mRNA transcription, the female adult germline produces transcripts that are not translated into proteins until embryogenesis of subsequent generation. As a consequence, translational control plays a central role in governing various germ cell decisions including the formation of primordial germ cells, self-renewal/differentiation decisions in the adult germline, onset of gametogenesis and oocyte maturation. Mechanistically, several common themes such as asymmetric localization of mRNAs, conserved RNA-binding proteins that control translation by 3' UTR binding, translational activation by the cytoplasmic elongation of the polyA tail and the assembly of mRNA-protein complexes called mRNPs have emerged from the studies on Caenorhabditis elegans, Xenopus and Drosophila. How mRNPs assemble, what influences their dynamics, and how a particular 3' UTR-binding protein turns on the translation of certain mRNAs while turning off other mRNAs at the same time and space are key challenges for future work.
Figures
References
-
- Arumugam K, MacNicol MC, Wang Y, Cragle CE, Tackett AJ, Hardy LL, MacNicol AM. Ringo/cyclin-dependent kinase and mitogen-activated protein kinase signaling pathways regulate the activity of the cell fate determinant Musashi to promote cell cycle re-entry in Xenopus oocytes. J Biol Chem. 2012b;287:10639–10649. - PMC - PubMed
-
- Austin J, Kimble J. glp-1 is required in the germ line for regulation of the decision between mitosis and meiosis in C. elegans. Cell. 1987;51:589–599. - PubMed
-
- Barnard DC, Ryan K, Manley JL, Richter JD. Symplekin and xGLD-2 are required for CPEB-mediated cytoplasmic polyadenylation. Cell. 2004;119:641–651. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
