

Cell death response to anti-mitotic drug treatment in cell culture, mouse tumor model and the clinic
- PMID: 28249963
- PMCID: PMC5557680
- DOI: 10.1530/ERC-17-0003
Cell death response to anti-mitotic drug treatment in cell culture, mouse tumor model and the clinic
Abstract
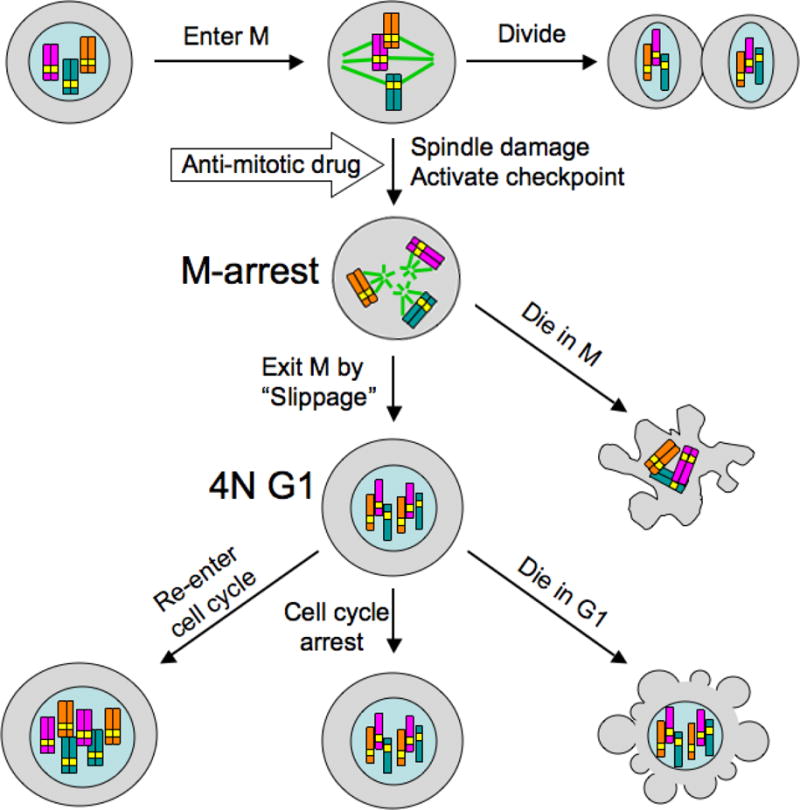
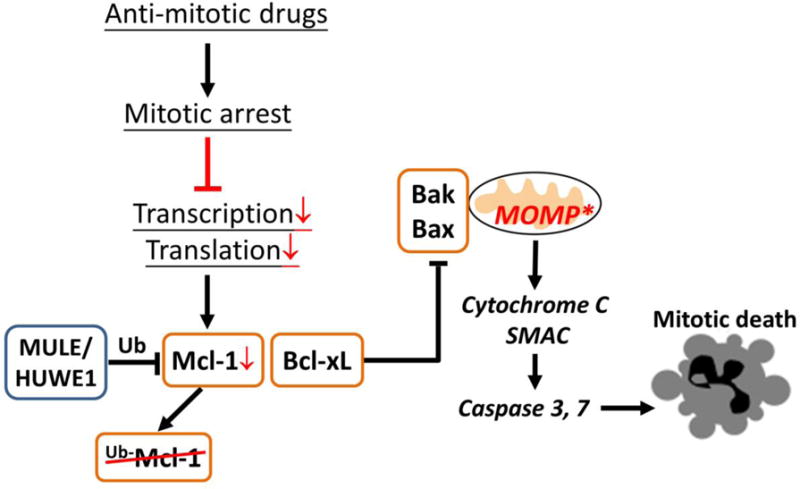
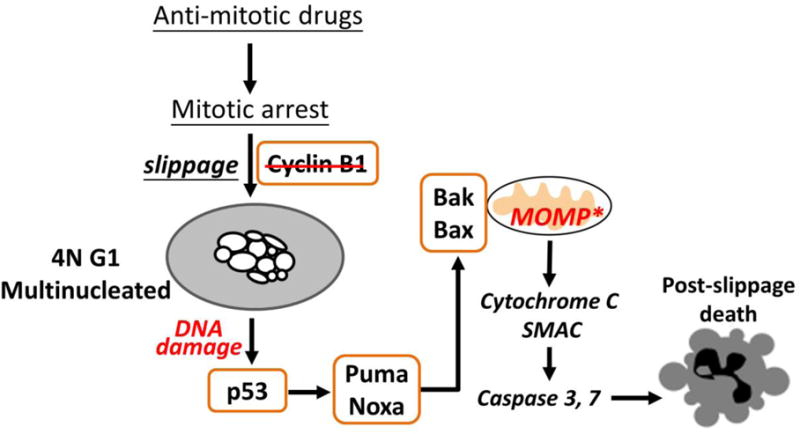
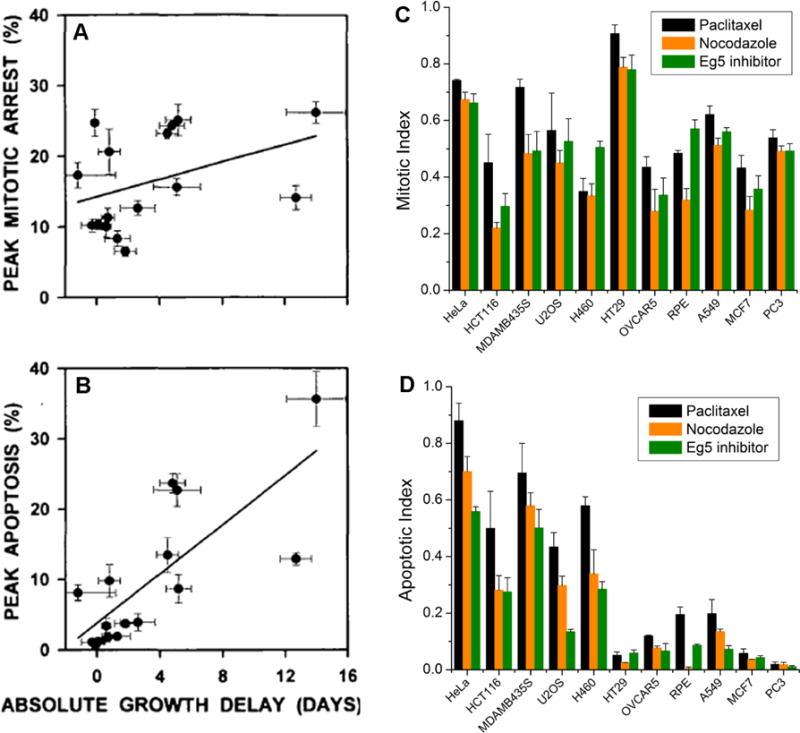
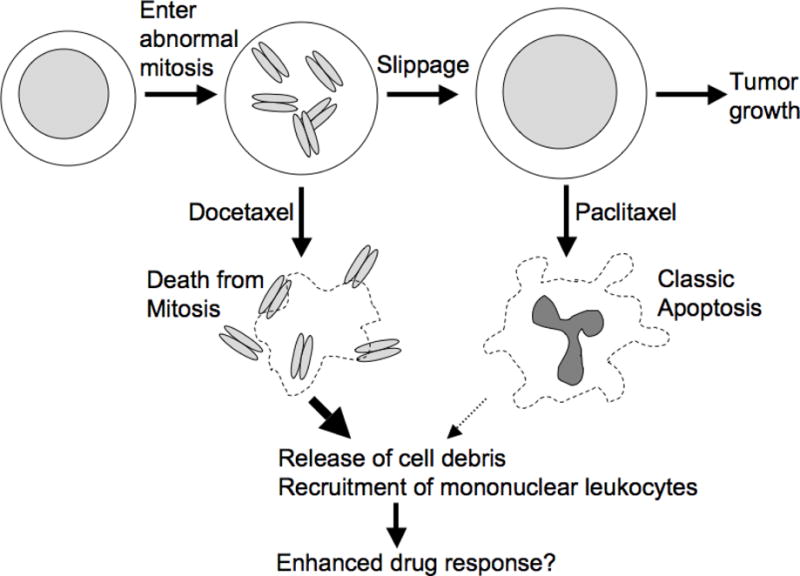
Anti-mitotic cancer drugs include classic microtubule-targeting drugs, such as taxanes and vinca alkaloids, and the newer spindle-targeting drugs, such as inhibitors of the motor protein; Kinesin-5 (aka KSP, Eg5, KIF11); and Aurora-A, Aurora-B and Polo-like kinases. Microtubule-targeting drugs are among the first line of chemotherapies for a wide spectrum of cancers, but patient responses vary greatly. We still lack understanding of how these drugs achieve a favorable therapeutic index, and why individual patient responses vary. Spindle-targeting drugs have so far shown disappointing results in the clinic, but it is possible that certain patients could benefit if we understand their mechanism of action better. Pre-clinical data from both cell culture and mouse tumor models showed that the cell death response is the most variable point of the drug action. Hence, in this review we focus on current mechanistic understanding of the cell death response to anti-mitotics. We first draw on extensive results from cell culture studies, and then cross-examine them with the more limited data from animal tumor models and the clinic. We end by discussing how cell type variation in cell death response might be harnessed to improve anti-mitotic chemotherapy by better patient stratification, new drug combinations and identification of novel targets for drug development.
Keywords: apoptosis; chemotherapy.
© 2017 Society for Endocrinology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Jordan MA, Wilson L. Microtubules as a target for anticancer drugs. Nat Rev Cancer. 2004;4:253–65. - PubMed
-
- Perez EA. Microtubule inhibitors: Differentiating tubulin-inhibiting agents based on mechanisms of action, clinical activity, and resistance. Mol Cancer Ther. 2009;8(8):2086–95. - PubMed
-
- Crown J, O’Leary M, Ooi WS. Docetaxel and paclitaxel in the treatment of breast cancer: a review of clinical experience. Oncologist. 2004;9(Suppl 2):24–32. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
