

Adult Muscle Formation Requires Drosophila Moleskin for Proliferation of Wing Disc-Associated Muscle Precursors
- PMID: 28249984
- PMCID: PMC5419470
- DOI: 10.1534/genetics.116.193813
Adult Muscle Formation Requires Drosophila Moleskin for Proliferation of Wing Disc-Associated Muscle Precursors
Abstract
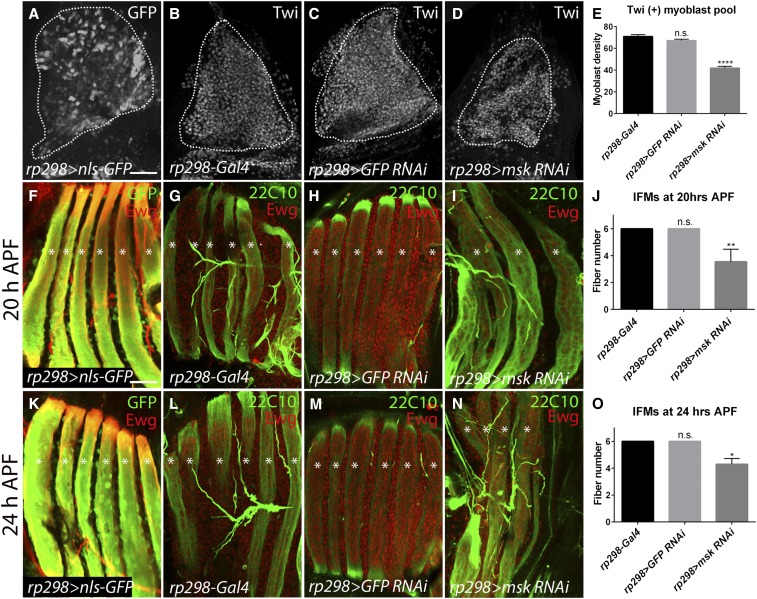
Adult muscle precursor (AMP) cells located in the notum of the larval wing disc undergo rapid amplification and eventual fusion to generate the Drosophila melanogaster indirect flight muscles (IFMs). Here we find that loss of Moleskin (Msk) function in these wing disc-associated myoblasts reduces the overall AMP pool size, resulting in the absence of IFM formation. This myoblast loss is due to a decrease in the AMP proliferative capacity and is independent of cell death. In contrast, disruption of Msk during pupal myoblast proliferation does not alter the AMP number, suggesting that Msk is specifically required for larval AMP proliferation. It has been previously shown that Wingless (Wg) signaling maintains expression of the Vestigial (Vg) transcription factor in proliferating myoblasts. However, other factors that influence Wg-mediated myoblast proliferation are largely unknown. Here we examine the interactions between Msk and the Wg pathway in regulation of the AMP pool size. We find that a myoblast-specific reduction of Msk results in the absence of Vg expression and a complete loss of the Wg pathway readout β-catenin/Armadillo (Arm). Moreover, msk RNA interference knockdown abolishes expression of the Wg target Ladybird (Lbe) in leg disc myoblasts. Collectively, our results provide strong evidence that Msk acts through the Wg signaling pathway to control myoblast pool size and muscle formation by regulating Arm stability or nuclear transport.
Keywords: Drosophila melanogaster; Moleskin; indirect flight muscles; proliferation.
Copyright © 2017 by the Genetics Society of America.
Figures







References
-
- Anant S., Roy S., VijayRaghavan K., 1998. Twist and Notch negatively regulate adult muscle differentiation in Drosophila. Development 125: 1361–1369. - PubMed
-
- Bate M., 1990. The embryonic development of larval muscles in Drosophila. Development 110: 791–804. - PubMed
-
- Bate M., Rushton E., Currie D. A., 1991. Cells with persistent twist expression are the embryonic precursors of adult muscles in Drosophila. Development 113: 79–89. - PubMed
-
- Bejsovec A., 2006. Flying at the head of the pack: Wnt biology in Drosophila. Oncogene 25: 7442–7449. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
