

Broad TCR repertoire and diverse structural solutions for recognition of an immunodominant CD8+ T cell epitope
- PMID: 28250417
- PMCID: PMC5383516
- DOI: 10.1038/nsmb.3383
Broad TCR repertoire and diverse structural solutions for recognition of an immunodominant CD8+ T cell epitope
Abstract
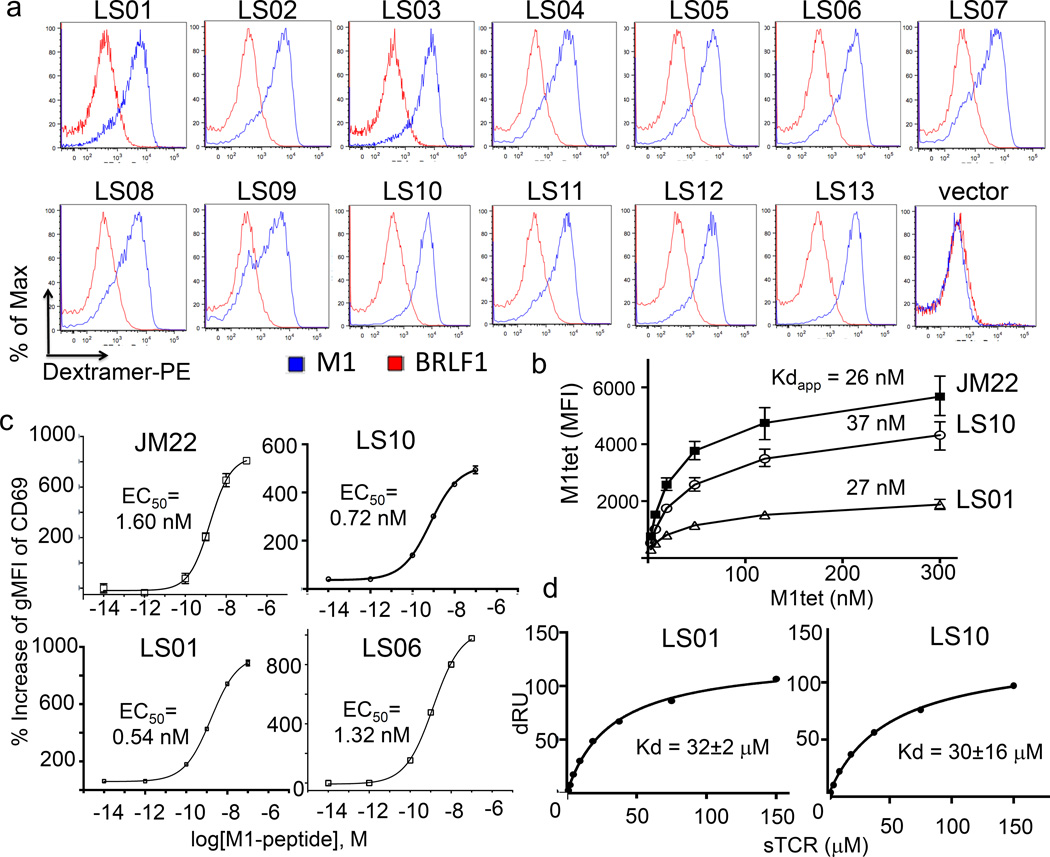
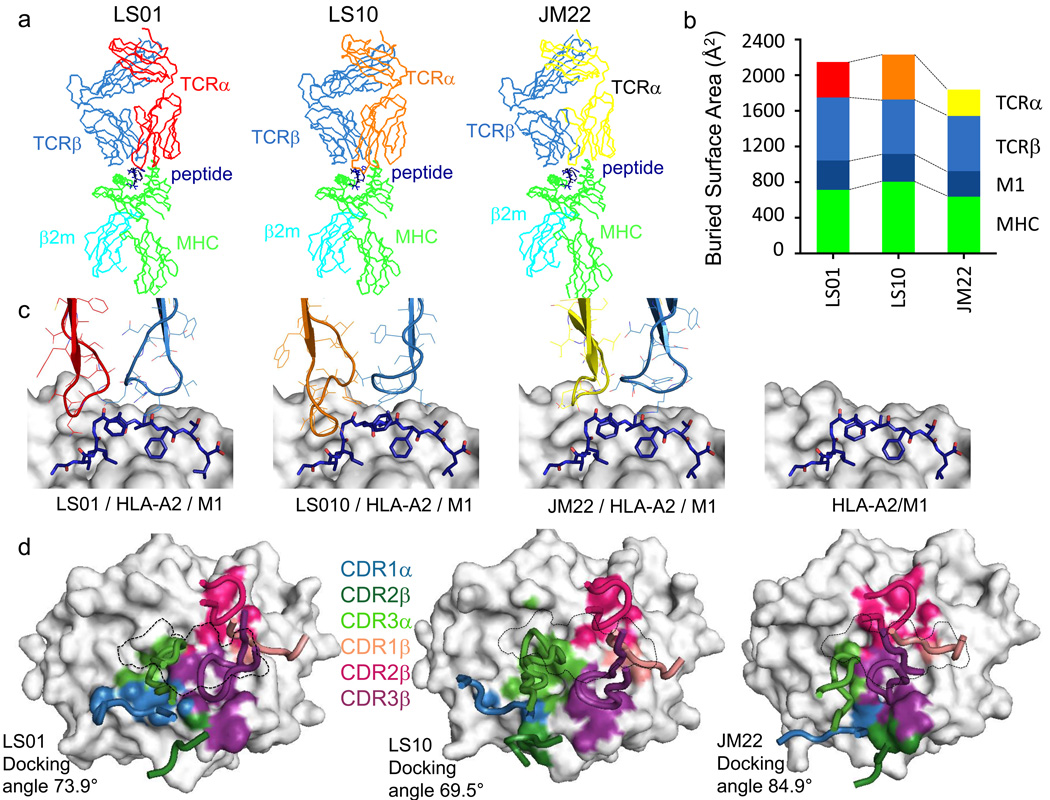
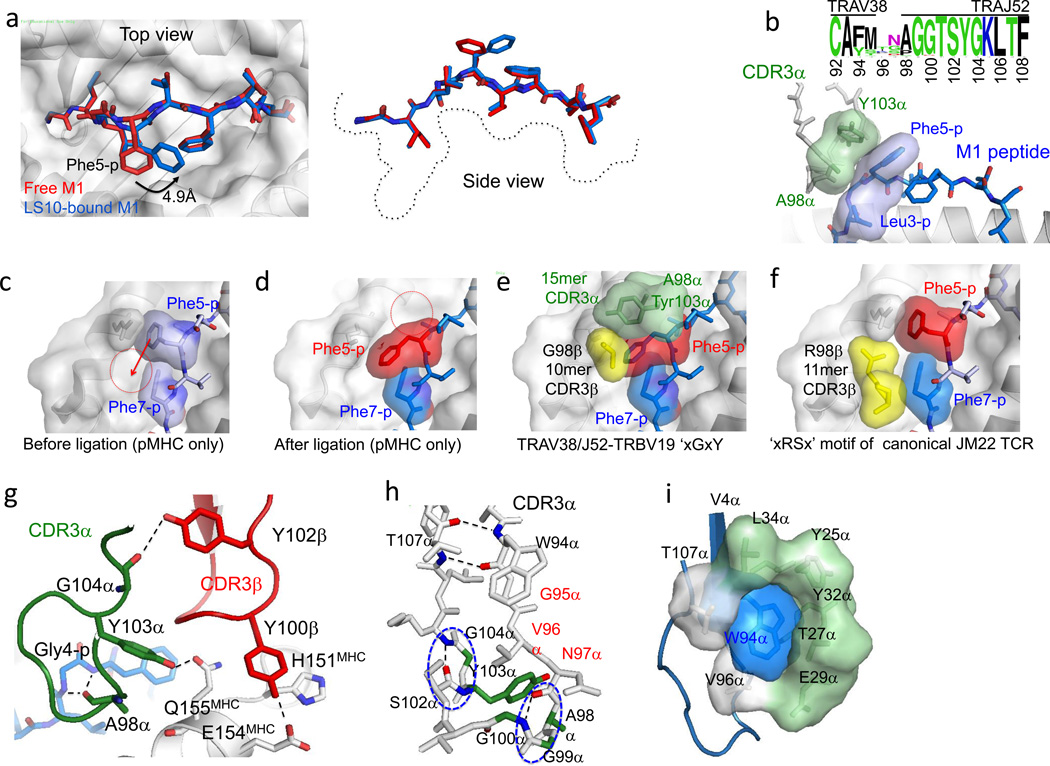
A keystone of antiviral immunity is CD8+ T cell recognition of viral peptides bound to MHC-I proteins. The recognition modes of individual T cell receptors (TCRs) have been studied in some detail, but the role of TCR variation in providing a robust response to viral antigens is unclear. The influenza M1 epitope is an immunodominant target of CD8+ T cells that help to control influenza in HLA-A2+ individuals. Here we show that CD8+ T cells use many distinct TCRs to recognize HLA-A2-M1, which enables the use of different structural solutions to the problem of specifically recognizing a relatively featureless peptide antigen. The vast majority of responding TCRs target a small cleft between HLA-A2 and the bound M1 peptide. These broad repertoires lead to plasticity in antigen recognition and protection against T cell clonal loss and viral escape.
Figures



References
-
- La Gruta NL, Turner SJ. T cell mediated immunity to influenza: mechanisms of viral control. Trends Immunol. 2014;35:396–402. - PubMed
-
- Tan ACL, et al. The design and proof of concept for a CD8+ T cell-based vaccine inducing cross-subtype protection against influenza A virus. Immunol. Cell Biol. 2013;91:96–104. - PubMed
-
- Gotch F, Rothbard J, Howland K, Townsend A, McMichael A. Cytotoxic T lymphocytes recognize a fragment of influenza virus matrix protein in association with HLA-A2. Nature. 1987;326:881–882. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
