

Tubulin acetylation protects long-lived microtubules against mechanical ageing
- PMID: 28250419
- PMCID: PMC5376231
- DOI: 10.1038/ncb3481
Tubulin acetylation protects long-lived microtubules against mechanical ageing
Abstract
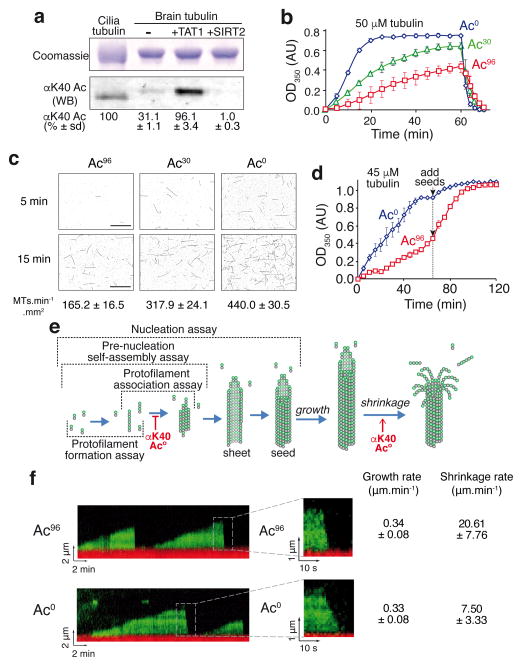
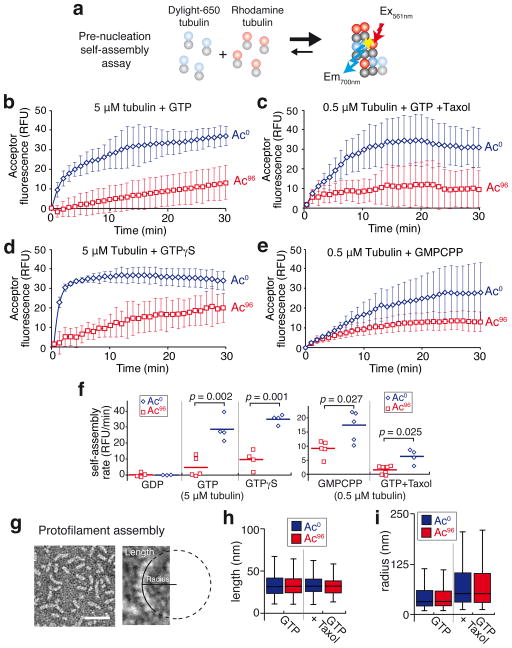
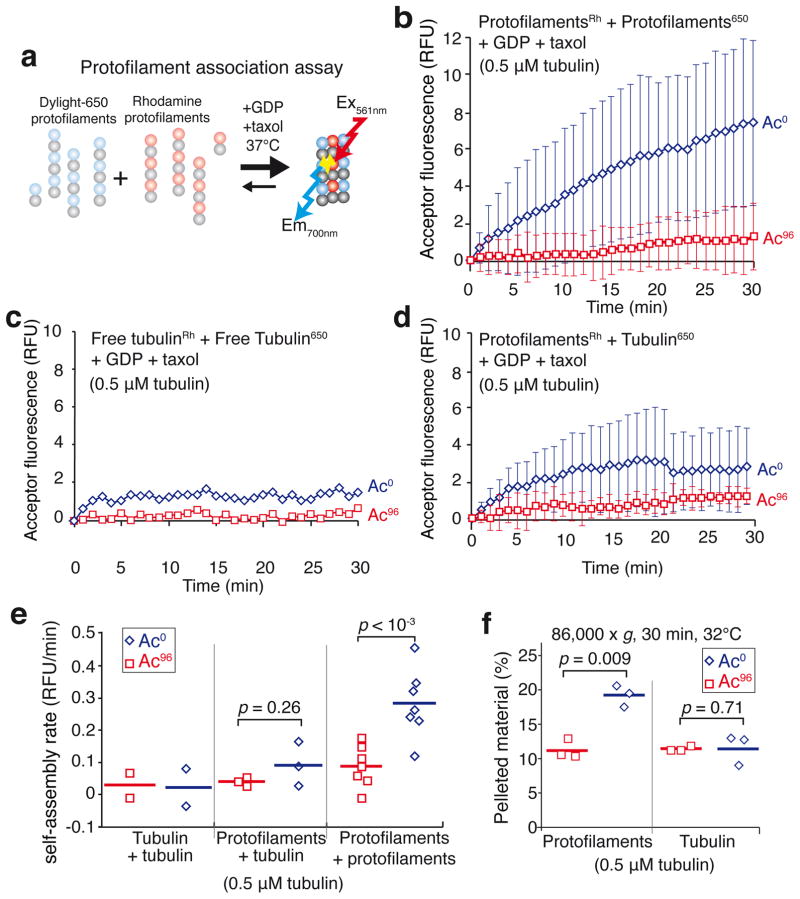
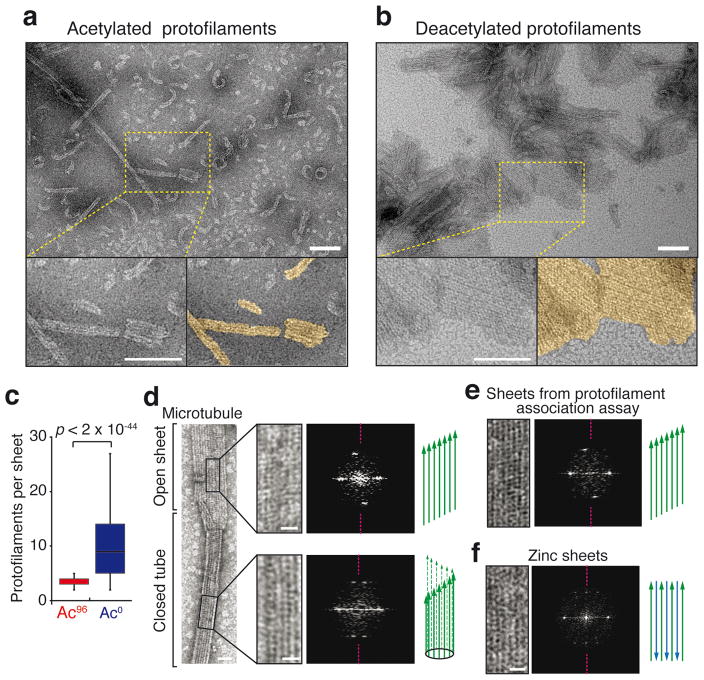
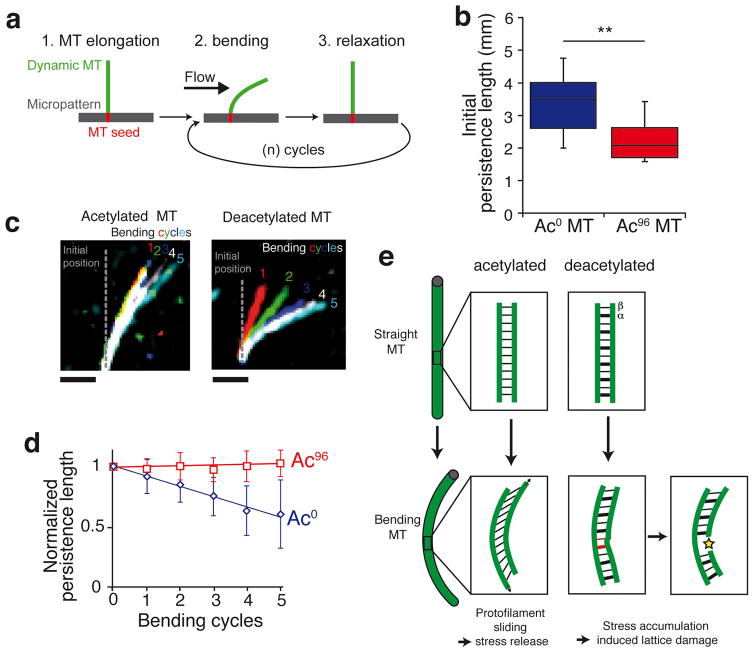
Long-lived microtubules endow the eukaryotic cell with long-range transport abilities. While long-lived microtubules are acetylated on Lys40 of α-tubulin (αK40), acetylation takes place after stabilization and does not protect against depolymerization. Instead, αK40 acetylation has been proposed to mechanically stabilize microtubules. Yet how modification of αK40, a residue exposed to the microtubule lumen and inaccessible to microtubule-associated proteins and motors, could affect microtubule mechanics remains an open question. Here we develop FRET-based assays that report on the lateral interactions between protofilaments and find that αK40 acetylation directly weakens inter-protofilament interactions. Congruently, αK40 acetylation affects two processes largely governed by inter-protofilament interactions, reducing the nucleation frequency and accelerating the shrinkage rate. Most relevant to the biological function of acetylation, microfluidics manipulations demonstrate that αK40 acetylation enhances flexibility and confers resilience against repeated mechanical stresses. Thus, unlike deacetylated microtubules that accumulate damage when subjected to repeated stresses, long-lived microtubules are protected from mechanical ageing through their acquisition of αK40 acetylation. In contrast to other tubulin post-translational modifications that act through microtubule-associated proteins, motors and severing enzymes, intraluminal acetylation directly tunes the compliance and resilience of microtubules.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
