

Allogeneic Adipose-Derived Mesenchymal Stromal Cells Ameliorate Experimental Autoimmune Encephalomyelitis by Regulating Self-Reactive T Cell Responses and Dendritic Cell Function
- PMID: 28250776
- PMCID: PMC5303870
- DOI: 10.1155/2017/2389753
Allogeneic Adipose-Derived Mesenchymal Stromal Cells Ameliorate Experimental Autoimmune Encephalomyelitis by Regulating Self-Reactive T Cell Responses and Dendritic Cell Function
Abstract
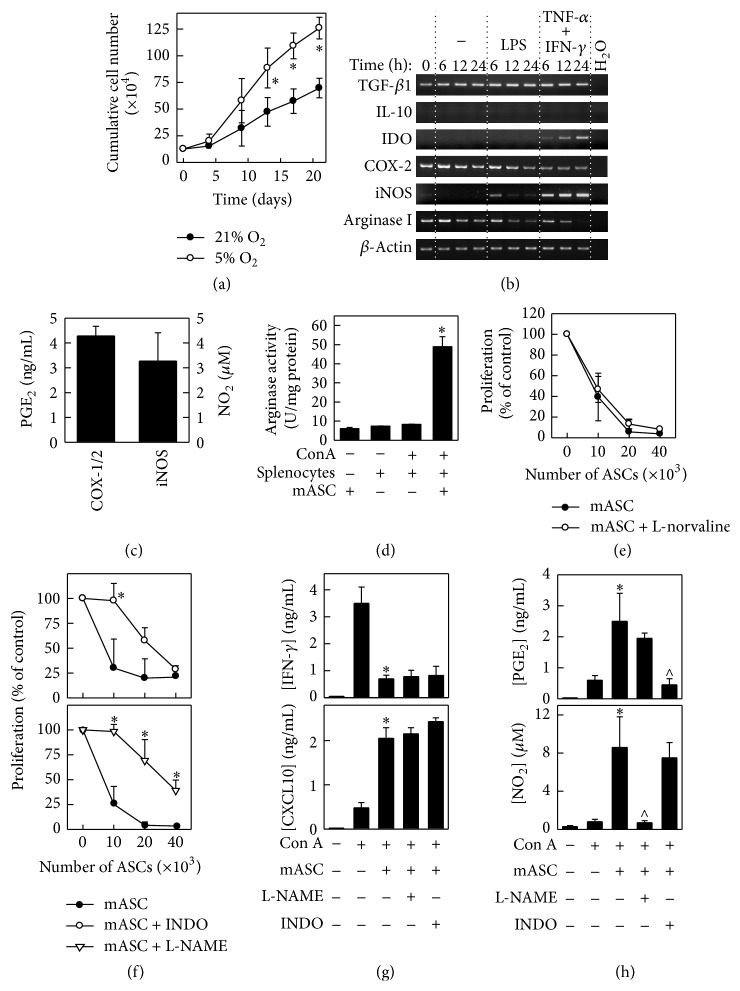
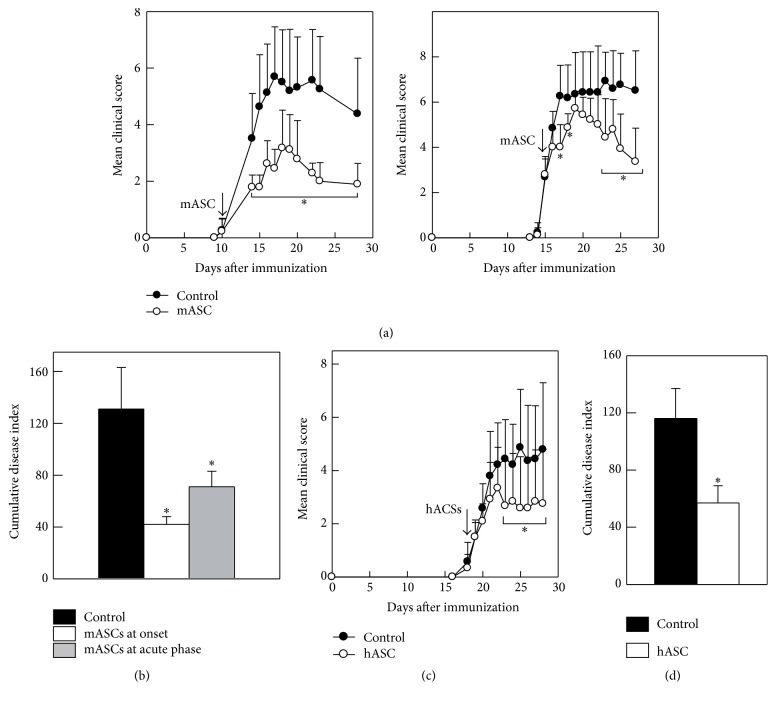
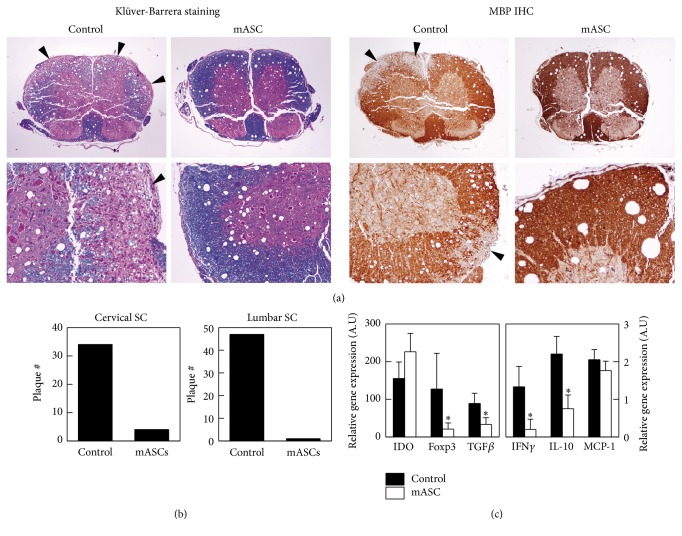
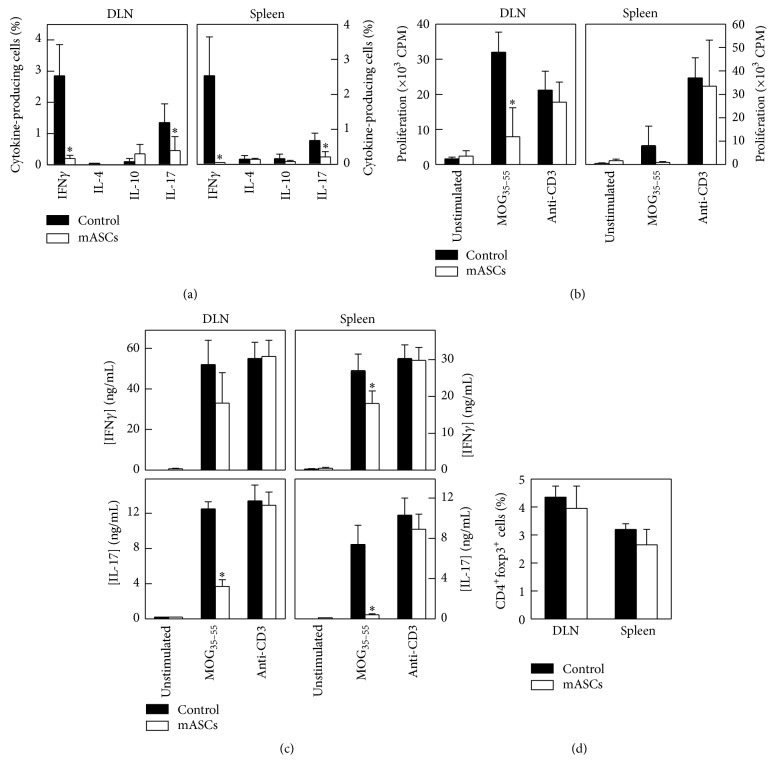
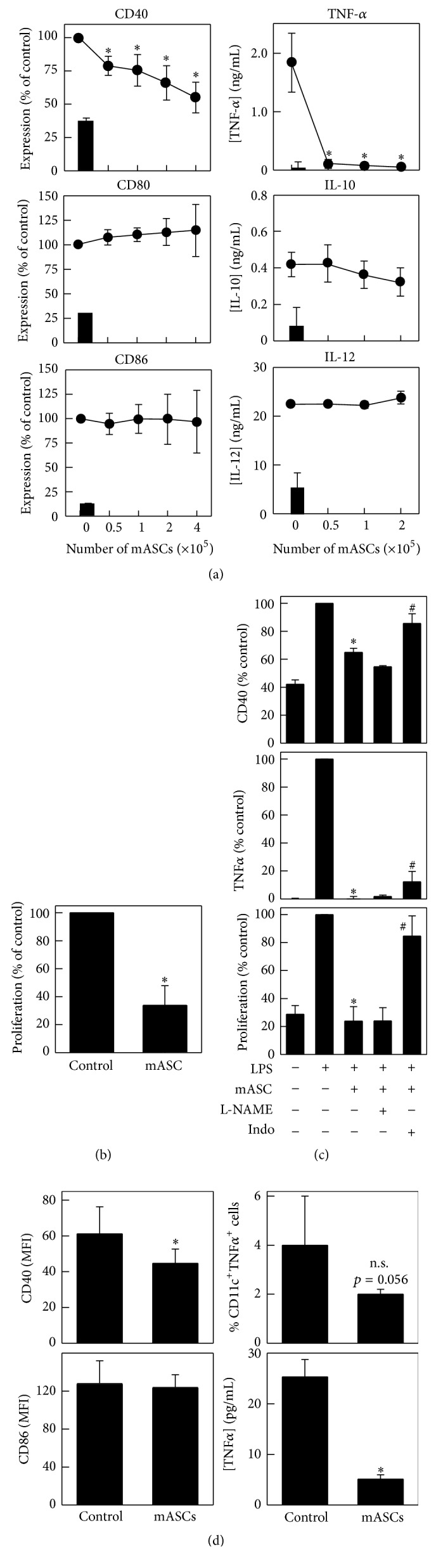
Multipotent mesenchymal stromal cells (MSCs) have emerged as a promising therapy for autoimmune diseases, including multiple sclerosis (MS). Administration of MSCs to MS patients has proven safe with signs of immunomodulation but their therapeutic efficacy remains low. The aim of the current study has been to further characterize the immunomodulatory mechanisms of adipose tissue-derived MSCs (ASCs) in vitro and in vivo using the EAE model of chronic brain inflammation in mice. We found that murine ASCs (mASCs) suppress T cell proliferation in vitro via inducible nitric oxide synthase (iNOS) and cyclooxygenase- (COX-) 1/2 activities. mASCs also prevented the lipopolysaccharide- (LPS-) induced maturation of dendritic cells (DCs) in vitro. The addition of the COX-1/2 inhibitor indomethacin, but not the iNOS inhibitor L-NAME, reversed the block in DC maturation implicating prostaglandin (PG) E2 in this process. In vivo, early administration of murine and human ASCs (hASCs) ameliorated myelin oligodendrocyte protein- (MOG35-55-) induced EAE in C57Bl/6 mice. Mechanistic studies showed that mASCs suppressed the function of autoantigen-specific T cells and also decreased the frequency of activated (CD11c+CD40high and CD11c+TNF-α+) DCs in draining lymph nodes (DLNs). In summary, these data suggest that mASCs reduce EAE severity, in part, through the impairment of DC and T cell function.
Conflict of interest statement
No competing financial interests exist
Figures
References
-
- Russell K. C., Phinney D. G., Lacey M. R., Barrilleaux B. L., Meyertholen K. E., O'Connor K. C. In vitro high-capacity assay to quantify the clonal heterogeneity in trilineage potential of mesenchymal stem cells reveals a complex hierarchy of lineage commitment. Stem Cells. 2010;28(4):788–798. doi: 10.1002/stem.312. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
