

Molecular Mechanisms Regulating Cell Fusion and Heterokaryon Formation in Filamentous Fungi
- PMID: 28256191
- PMCID: PMC11687462
- DOI: 10.1128/microbiolspec.FUNK-0015-2016
Molecular Mechanisms Regulating Cell Fusion and Heterokaryon Formation in Filamentous Fungi
Abstract
For the majority of fungal species, the somatic body of an individual is a network of interconnected cells sharing a common cytoplasm and organelles. This syncytial organization contributes to an efficient distribution of resources, energy, and biochemical signals. Cell fusion is a fundamental process for fungal development, colony establishment, and habitat exploitation and can occur between hyphal cells of an individual colony or between colonies of genetically distinct individuals. One outcome of cell fusion is the establishment of a stable heterokaryon, culminating in benefits for each individual via shared resources or being of critical importance for the sexual or parasexual cycle of many fungal species. However, a second outcome of cell fusion between genetically distinct strains is formation of unstable heterokaryons and the induction of a programmed cell death reaction in the heterokaryotic cells. This reaction of nonself rejection, which is termed heterokaryon (or vegetative) incompatibility, is widespread in the fungal kingdom and acts as a defense mechanism against genome exploitation and mycoparasitism. Here, we review the currently identified molecular players involved in the process of somatic cell fusion and its regulation in filamentous fungi. Thereafter, we summarize the knowledge of the molecular determinants and mechanism of heterokaryon incompatibility and place this phenomenon in the broader context of biotropic interactions and immunity.
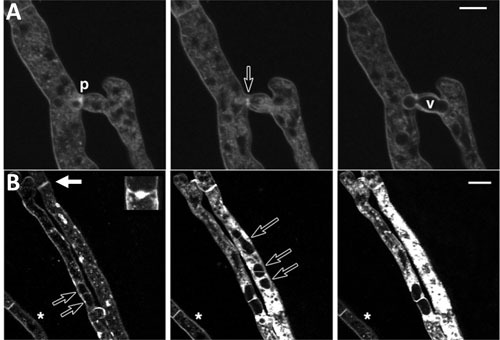
Figures






References
-
- Read ND, Goryachev AB, Lichius A. 2012. The mechanistic basis of self-fusion between conidial anastomosis tubes during fungal colony initiation. Fungal Biol Rev 26:1–11. 10.1016/j.fbr.2012.02.003 - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
