

Toxoplasma growth in vitro is dependent on exogenous tyrosine and is independent of AAH2 even in tyrosine-limiting conditions
- PMID: 28257757
- PMCID: PMC5423395
- DOI: 10.1016/j.exppara.2017.02.018
Toxoplasma growth in vitro is dependent on exogenous tyrosine and is independent of AAH2 even in tyrosine-limiting conditions
Abstract
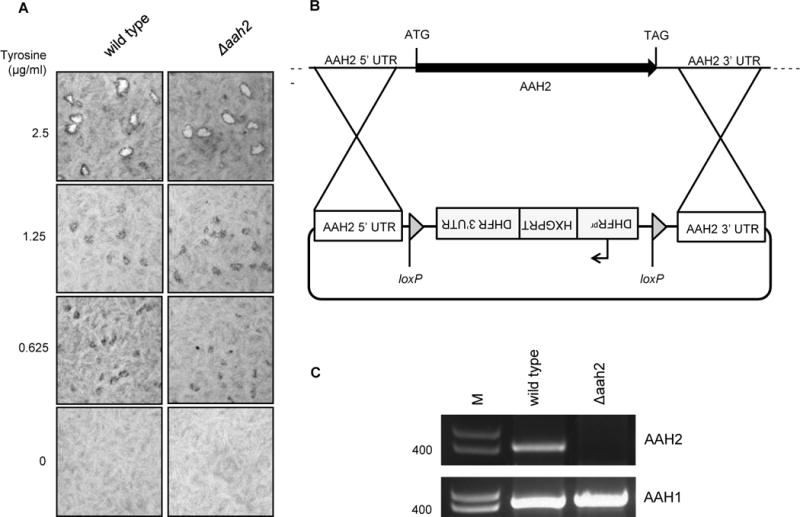
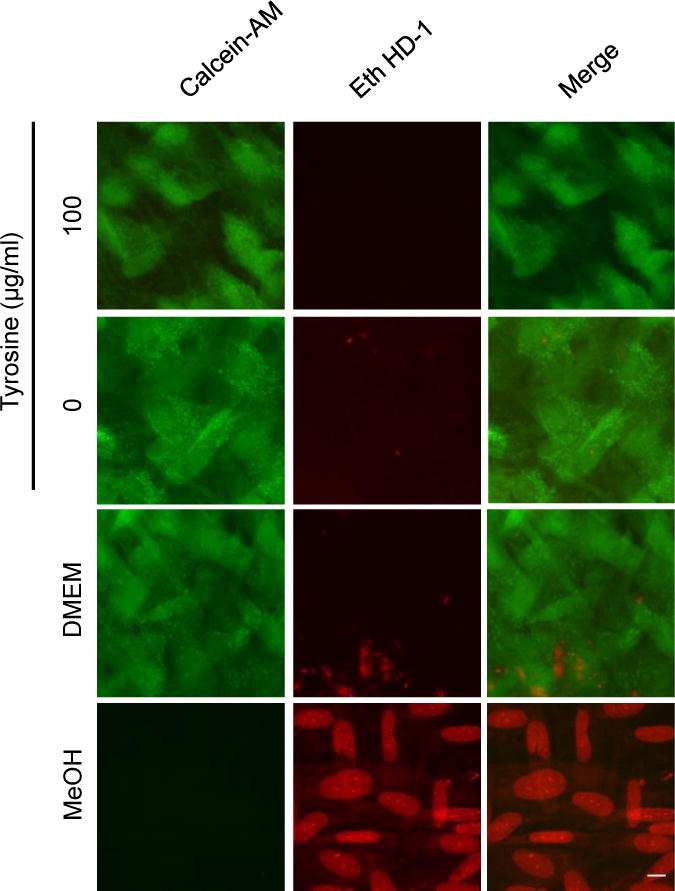
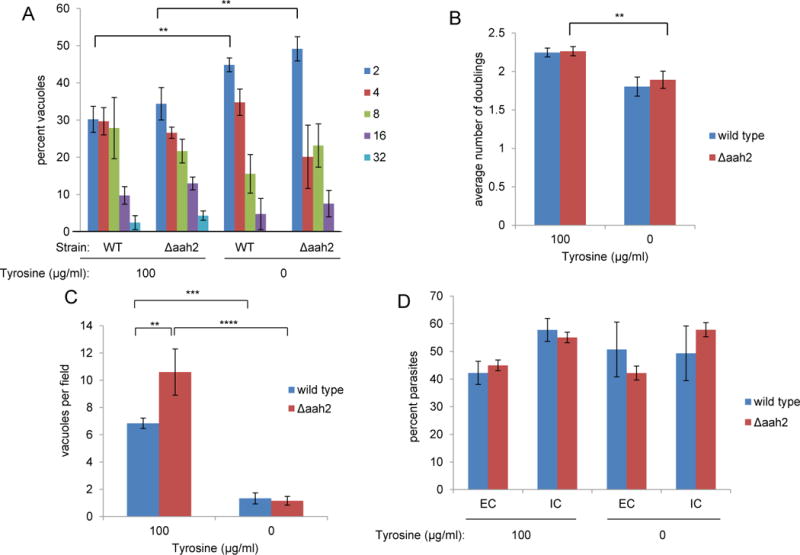
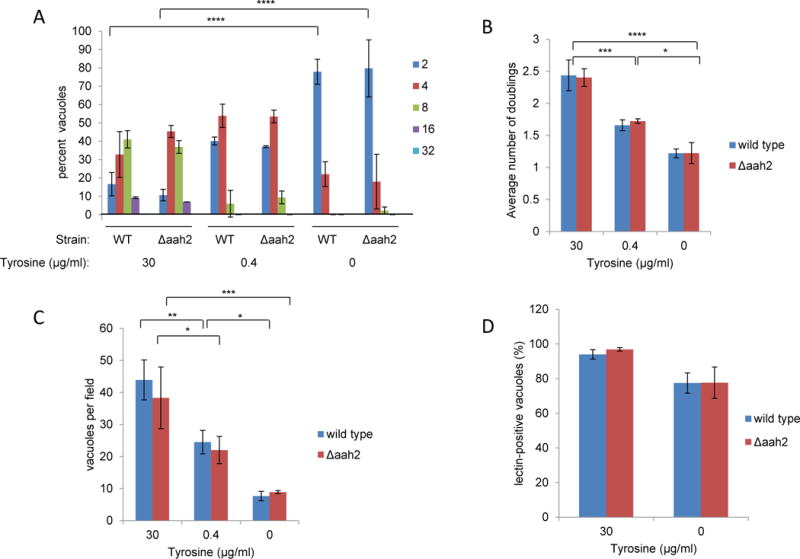
Toxoplasma gondii is an obligate intracellular parasite capable of infecting virtually all nucleated cell types in almost all warm-blooded animals. Interestingly, Toxoplasma has a relatively full repertoire of amino acid biosynthetic machinery, perhaps reflecting its broad host range and, consequently, its need to adapt to a wide array of amino acid resources. Although Toxoplasma has been shown to be auxotrophic for tryptophan and arginine, it has not previously been determined if Toxoplasma is also auxotrophic for tyrosine. Toxoplasma tachyzoites and bradyzoites were recently found to express an amino acid hydroxylase (AAH2) that is capable of synthesizing tyrosine and dihydroxyphenylalanine (DOPA) from phenylalanine; however, the role of AAH2 in tachyzoite and bradyzoite infection has not yet been identified. To determine if Toxoplasma requires exogenous tyrosine for growth, we performed growth assays on tachyzoites and bradyzoites in nutrient-rich media titrated with varying amounts of tyrosine. We found that Toxoplasma tachyzoites form significantly smaller plaques in tyrosine-limiting media in a dose-dependent manner and that this phenotype is not affected by deletion of TgAAH2. To determine if bradyzoites require exogenous tyrosine for growth, we induced differentiation from tachyzoites in vitro in tyrosine-limiting media and found that replication and vacuole number are all decreased in tyrosine-deficient media. Importantly, culture of confluent human fibroblasts in tyrosine-deficient media does not affect their viability, indicating that, at least in vitro, the need for tyrosine is at the level of Toxoplasma, not the host cell supporting its growth.
Keywords: Amino acid hydroxylase; Apicomplexan; Metabolism.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Toxoplasma gondii Cyclic AMP-Dependent Protein Kinase Subunit 3 Is Involved in the Switch from Tachyzoite to Bradyzoite Development.mBio. 2016 May 31;7(3):e00755-16. doi: 10.1128/mBio.00755-16. mBio. 2016. PMID: 27247232 Free PMC article.
-
Toxoplasma gondii lacks the enzymes required for de novo arginine biosynthesis and arginine starvation triggers cyst formation.Int J Parasitol. 2004 Mar 9;34(3):323-31. doi: 10.1016/j.ijpara.2003.12.001. Int J Parasitol. 2004. PMID: 15003493
-
Primary skeletal muscle cells trigger spontaneous Toxoplasma gondii tachyzoite-to-bradyzoite conversion at higher rates than fibroblasts.Int J Med Microbiol. 2009 Jun;299(5):381-8. doi: 10.1016/j.ijmm.2008.10.002. Epub 2008 Dec 18. Int J Med Microbiol. 2009. PMID: 19097936
-
The determinants regulating Toxoplasma gondii bradyzoite development.Front Microbiol. 2022 Nov 11;13:1027073. doi: 10.3389/fmicb.2022.1027073. eCollection 2022. Front Microbiol. 2022. PMID: 36439853 Free PMC article. Review.
-
Stress-related and spontaneous stage differentiation of Toxoplasma gondii.Mol Biosyst. 2008 Aug;4(8):824-34. doi: 10.1039/b800520f. Epub 2008 Jun 2. Mol Biosyst. 2008. PMID: 18633484 Review.
Cited by
-
Dual metabolomic profiling uncovers Toxoplasma manipulation of the host metabolome and the discovery of a novel parasite metabolic capability.PLoS Pathog. 2020 Apr 7;16(4):e1008432. doi: 10.1371/journal.ppat.1008432. eCollection 2020 Apr. PLoS Pathog. 2020. PMID: 32255806 Free PMC article.
-
Amino Acid Metabolism in Apicomplexan Parasites.Metabolites. 2021 Jan 20;11(2):61. doi: 10.3390/metabo11020061. Metabolites. 2021. PMID: 33498308 Free PMC article. Review.
-
Metabolic adaptability and nutrient scavenging in Toxoplasma gondii: insights from ingestion pathway-deficient mutants.mSphere. 2025 Apr 29;10(4):e0101124. doi: 10.1128/msphere.01011-24. Epub 2025 Apr 2. mSphere. 2025. PMID: 40172222 Free PMC article.
-
4-Arylthiosemicarbazide Derivatives as Toxoplasmic Aromatic Amino Acid Hydroxylase Inhibitors and Anti-inflammatory Agents.Int J Mol Sci. 2022 Mar 16;23(6):3213. doi: 10.3390/ijms23063213. Int J Mol Sci. 2022. PMID: 35328634 Free PMC article.
-
The Bradyzoite: A Key Developmental Stage for the Persistence and Pathogenesis of Toxoplasmosis.Pathogens. 2020 Mar 21;9(3):234. doi: 10.3390/pathogens9030234. Pathogens. 2020. PMID: 32245165 Free PMC article. Review.
References
-
- Dubey JP. Advances in the life cycle of Toxoplasma gondii. Int J Parasitol. 1998;28(7):1019–24. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
