

Activation Mechanism and Cellular Localization of Membrane-Anchored Alginate Polymerase in Pseudomonas aeruginosa
- PMID: 28258142
- PMCID: PMC5394334
- DOI: 10.1128/AEM.03499-16
Activation Mechanism and Cellular Localization of Membrane-Anchored Alginate Polymerase in Pseudomonas aeruginosa
Abstract
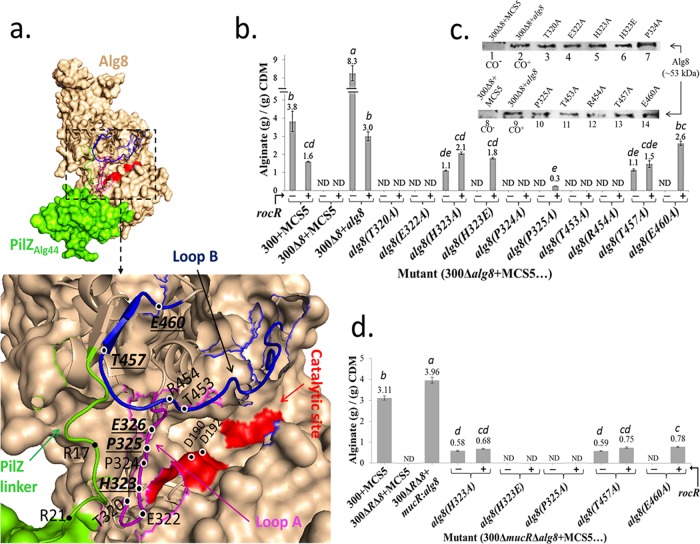
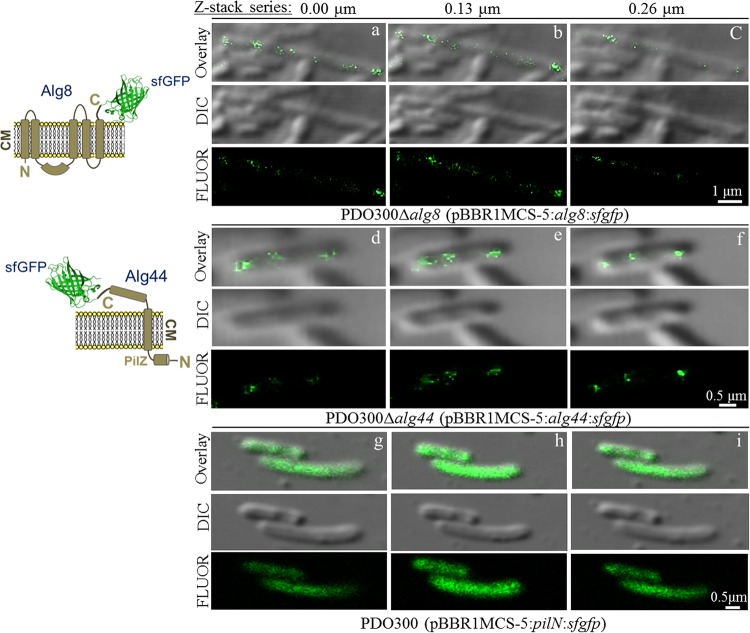
The exopolysaccharide alginate, produced by the opportunistic human pathogen Pseudomonas aeruginosa, confers a survival advantage to the bacterium by contributing to the formation of characteristic biofilms during infection. Membrane-anchored proteins Alg8 (catalytic subunit) and Alg44 (copolymerase) constitute the alginate polymerase that is being activated by the second messenger molecule bis-(3', 5')-cyclic dimeric GMP (c-di-GMP), but the mechanism of activation remains elusive. To shed light on the c-di-GMP-mediated activation of alginate polymerization in vivo, an in silico structural model of Alg8 fused to the c-di-GMP binding PilZ domain informed by the structure of cellulose synthase, BcsA, was developed. This structural model was probed by site-specific mutagenesis and different cellular levels of c-di-GMP. Results suggested that c-di-GMP-mediated activation of alginate polymerization involves amino acids residing at two loops, including H323 (loop A) and T457 and E460 (loop B), surrounding the catalytic site in the predicted model. The activities of the respective Alg8 variants suggested that c-di-GMP-mediated control of substrate access to the catalytic site of Alg8 is dissimilar to the known activation mechanism of BcsA. Alg8 variants responded differently to various c-di-GMP levels, while MucR imparted c-di-GMP for activation of alginate polymerase. Furthermore, we showed that Alg44 copolymerase constituted a stable dimer, with its periplasmic domains required for protein localization and alginate polymerization and modification. Superfolder green fluorescent protein (GFP) fusions of Alg8 and Alg44 showed a nonuniform, punctate, and patchy arrangement of both proteins surrounding the cell. Overall, this study provides insights into the c-di-GMP-mediated activation of alginate polymerization while assigning functional roles to Alg8 and Alg44, including their subcellular localization and distribution.IMPORTANCE The exopolysaccharide alginate is an important biofilm component of the opportunistic human pathogen P. aeruginosa and the principal cause of the mucoid phenotype that is the hallmark of chronic infections of cystic fibrosis patients. The production of alginate is mediated by interacting membrane proteins Alg8 and Alg44, while their activity is posttranslationally regulated by the second messenger c-di-GMP, a well-known regulator of the synthesis of a range of other exopolysaccharides in bacteria. This study provides new insights into the unknown activation mechanism of alginate polymerization by c-di-GMP. Experimental evidence that the activation of alginate polymerization requires the engagement of specific amino acid residues residing at the catalytic domain of Alg8 glycosyltransferase was obtained, and these residues are proposed to exert an allosteric effect on the PilZAlg44 domain upon c-di-GMP binding. This mechanism is dissimilar to the proposed mechanism of the autoinhibition of cellulose polymerization imposed by salt bridge formation between amino acid residues and released upon c-di-GMP binding, leading to activation of polymerization. On the other hand, conserved amino acid residues in the periplasmic domain of Alg44 were found to be involved in alginate polymerization as well as modification events, i.e., acetylation and epimerization. Due to the critical role of c-di-GMP in the regulation of many biological processes, particularly the motility-sessility switch and also the emergence of persisting mucoid phenotypes, these results aid to reach a better understanding of biofilm-associated regulatory networks and c-di-GMP signaling and might assist the development of inhibitory drugs.
Keywords: Pseudomonas aeruginosa; alginate; polymerases.
Copyright © 2017 American Society for Microbiology.
Figures

Similar articles
-
Alginate Polymerization and Modification Are Linked in Pseudomonas aeruginosa.mBio. 2015 May 12;6(3):e00453-15. doi: 10.1128/mBio.00453-15. mBio. 2015. PMID: 25968647 Free PMC article.
-
Membrane topology and roles of Pseudomonas aeruginosa Alg8 and Alg44 in alginate polymerization.Microbiology (Reading). 2008 Jun;154(Pt 6):1605-1615. doi: 10.1099/mic.0.2007/015305-0. Microbiology (Reading). 2008. PMID: 18524915 Free PMC article.
-
The second messenger bis-(3'-5')-cyclic-GMP and its PilZ domain-containing receptor Alg44 are required for alginate biosynthesis in Pseudomonas aeruginosa.Mol Microbiol. 2007 Aug;65(4):876-95. doi: 10.1111/j.1365-2958.2007.05817.x. Epub 2007 Jul 21. Mol Microbiol. 2007. PMID: 17645452
-
Matrix exopolysaccharides; the sticky side of biofilm formation.FEMS Microbiol Lett. 2017 Jul 6;364(13):fnx120. doi: 10.1093/femsle/fnx120. FEMS Microbiol Lett. 2017. PMID: 28605431 Free PMC article. Review.
-
Emerging paradigms for PilZ domain-mediated C-di-GMP signaling.Biochem Soc Trans. 2019 Feb 28;47(1):381-388. doi: 10.1042/BST20180543. Epub 2019 Feb 1. Biochem Soc Trans. 2019. PMID: 30710060 Review.
Cited by
-
Transcriptome sequencing of Saccharina japonica sporophytes during whole developmental periods reveals regulatory networks underlying alginate and mannitol biosynthesis.BMC Genomics. 2019 Dec 12;20(1):975. doi: 10.1186/s12864-019-6366-x. BMC Genomics. 2019. PMID: 31830918 Free PMC article.
-
Harnessing Metabolic Regulation to Increase Hfq-Dependent Antibiotic Susceptibility in Pseudomonas aeruginosa.Front Microbiol. 2018 Nov 9;9:2709. doi: 10.3389/fmicb.2018.02709. eCollection 2018. Front Microbiol. 2018. PMID: 30473687 Free PMC article.
-
Analysis of Sanitizer Rotation on the Susceptibility, Biofilm Forming Ability and Caco-2 Cell Adhesion and Invasion of Listeria.Pathogens. 2022 Aug 24;11(9):961. doi: 10.3390/pathogens11090961. Pathogens. 2022. PMID: 36145393 Free PMC article.
-
The TPR domain of PgaA is a multifunctional scaffold that binds PNAG and modulates PgaB-dependent polymer processing.PLoS Pathog. 2022 Aug 5;18(8):e1010750. doi: 10.1371/journal.ppat.1010750. eCollection 2022 Aug. PLoS Pathog. 2022. PMID: 35930610 Free PMC article.
-
Regulation of Biofilm Exopolysaccharide Production by Cyclic Di-Guanosine Monophosphate.Front Microbiol. 2021 Sep 10;12:730980. doi: 10.3389/fmicb.2021.730980. eCollection 2021. Front Microbiol. 2021. PMID: 34566936 Free PMC article. Review.
References
-
- May TB, Shinabarger D, Maharaj R, Kato J, Chu L, DeVault JD, Roychoudhury S, Zielinski NA, Berry A, Rothmel RK. 1991. Alginate synthesis by Pseudomonas aeruginosa: a key pathogenic factor in chronic pulmonary infections of cystic fibrosis patients. Clin Microbiol Rev 4:191–206. doi:10.1128/CMR.4.2.191. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
