

Annealing of Complementary DNA Sequences During Double-Strand Break Repair in Drosophila Is Mediated by the Ortholog of SMARCAL1
- PMID: 28258182
- PMCID: PMC5419489
- DOI: 10.1534/genetics.117.200238
Annealing of Complementary DNA Sequences During Double-Strand Break Repair in Drosophila Is Mediated by the Ortholog of SMARCAL1
Abstract
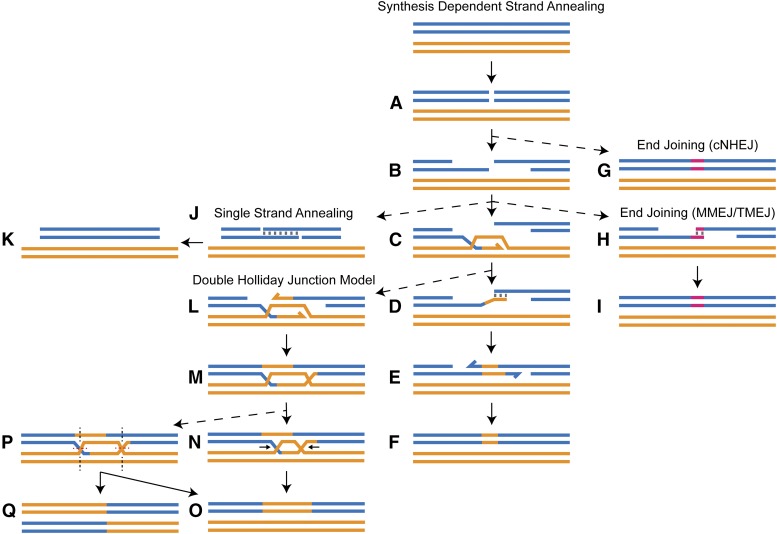
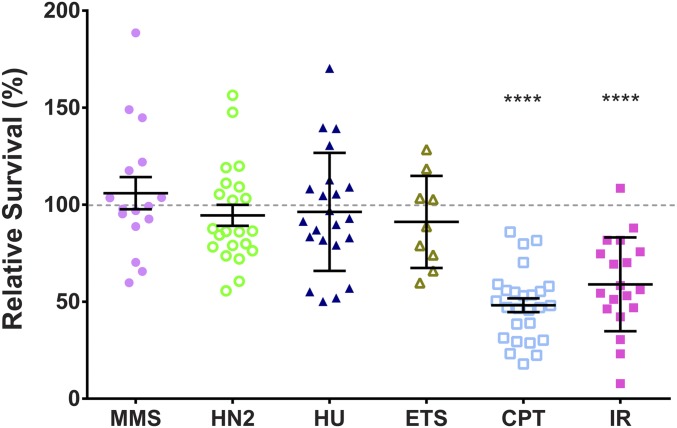
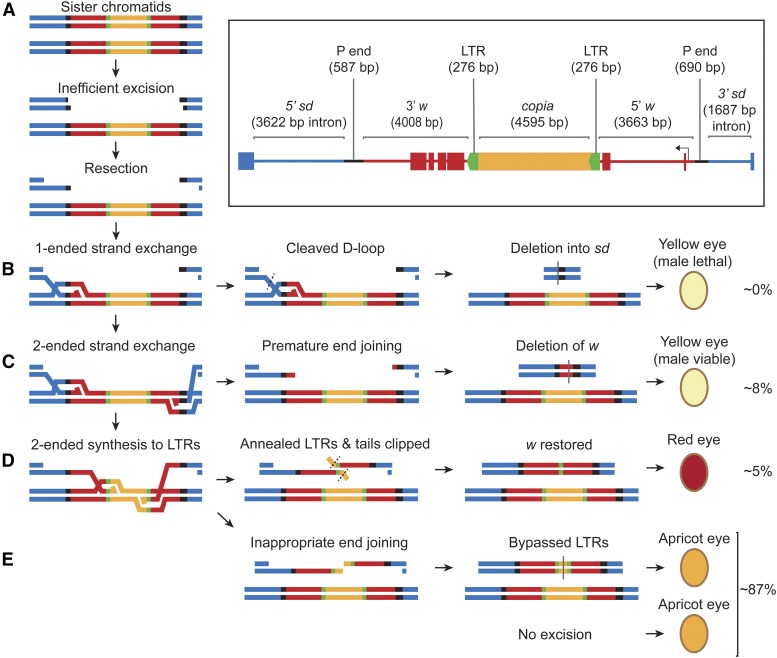
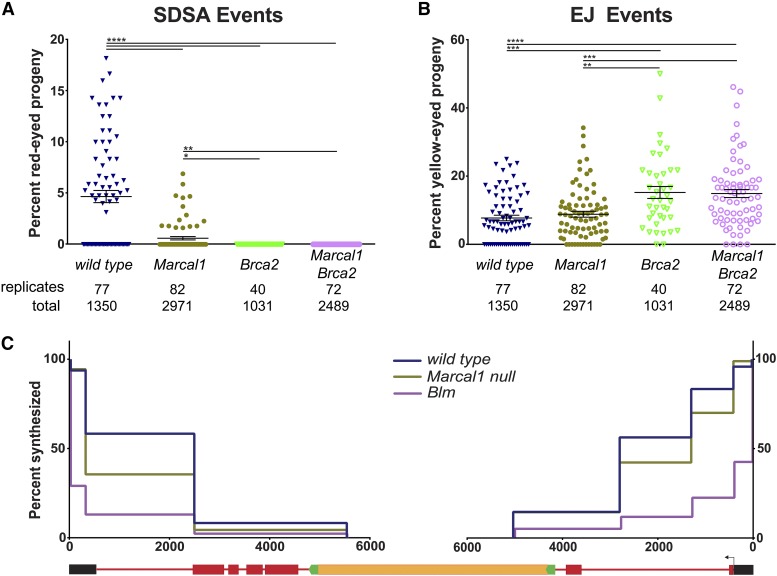
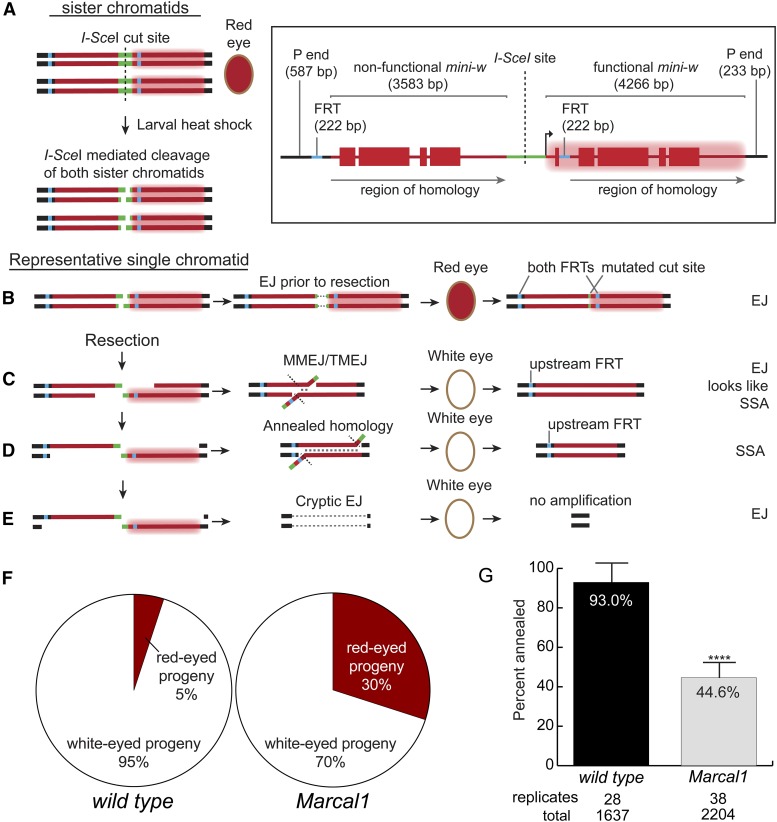
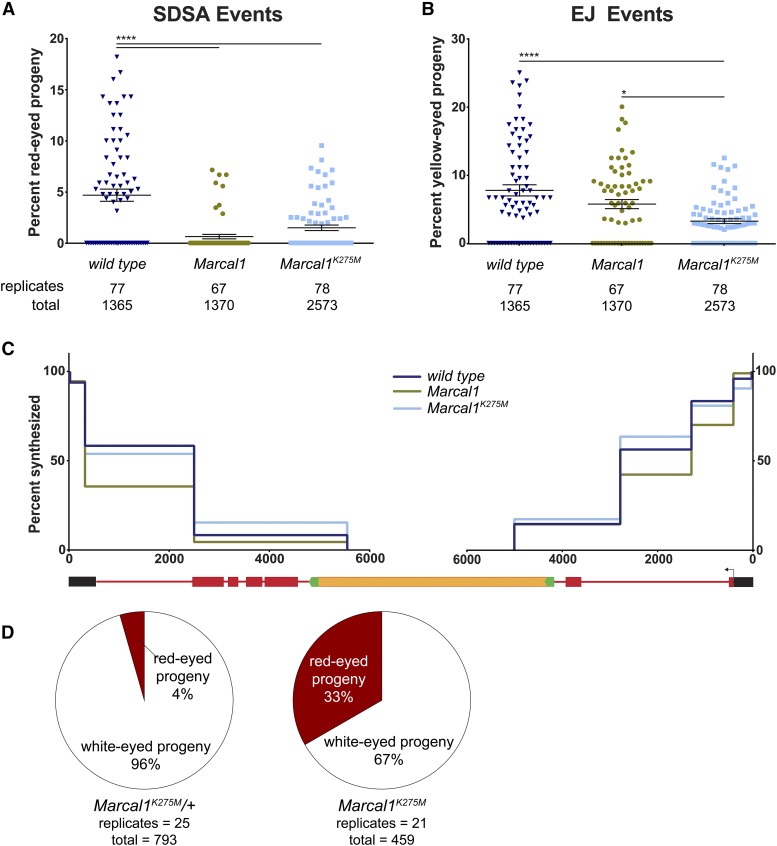
DNA double-strand breaks (DSBs) pose a serious threat to genomic integrity. If unrepaired, they can lead to chromosome fragmentation and cell death. If repaired incorrectly, they can cause mutations and chromosome rearrangements. DSBs are repaired using end-joining or homology-directed repair strategies, with the predominant form of homology-directed repair being synthesis-dependent strand annealing (SDSA). SDSA is the first defense against genomic rearrangements and information loss during DSB repair, making it a vital component of cell health and an attractive target for chemotherapeutic development. SDSA has also been proposed to be the primary mechanism for integration of large insertions during genome editing with CRISPR/Cas9. Despite the central role for SDSA in genome stability, little is known about the defining step: annealing. We hypothesized that annealing during SDSA is performed by the annealing helicase SMARCAL1, which can anneal RPA-coated single DNA strands during replication-associated DNA damage repair. We used unique genetic tools in Drosophila melanogaster to test whether the fly ortholog of SMARCAL1, Marcal1, mediates annealing during SDSA. Repair that requires annealing is significantly reduced in Marcal1 null mutants in both synthesis-dependent and synthesis-independent (single-strand annealing) assays. Elimination of the ATP-binding activity of Marcal1 also reduced annealing-dependent repair, suggesting that the annealing activity requires translocation along DNA. Unlike the null mutant, however, the ATP-binding defect mutant showed reduced end joining, shedding light on the interaction between SDSA and end-joining pathways.
Keywords: DSB repair; Drosophila; SDSA; SMARCAL1; homologous recombination.
Copyright © 2017 by the Genetics Society of America.
Figures
Similar articles
-
Single-Strand Annealing Plays a Major Role in Double-Strand DNA Break Repair following CRISPR-Cas9 Cleavage in Leishmania.mSphere. 2019 Aug 21;4(4):e00408-19. doi: 10.1128/mSphere.00408-19. mSphere. 2019. PMID: 31434745 Free PMC article.
-
Pathways and assays for DNA double-strand break repair by homologous recombination.Acta Biochim Biophys Sin (Shanghai). 2019 Sep 6;51(9):879-889. doi: 10.1093/abbs/gmz076. Acta Biochim Biophys Sin (Shanghai). 2019. PMID: 31294447 Review.
-
The Role of Drosophila CtIP in Homology-Directed Repair of DNA Double-Strand Breaks.Genes (Basel). 2021 Sep 16;12(9):1430. doi: 10.3390/genes12091430. Genes (Basel). 2021. PMID: 34573412 Free PMC article.
-
Smarcal1 promotes double-strand-break repair by nonhomologous end-joining.Nucleic Acids Res. 2015 Jul 27;43(13):6359-72. doi: 10.1093/nar/gkv621. Epub 2015 Jun 18. Nucleic Acids Res. 2015. PMID: 26089390 Free PMC article.
-
[SMARCAL1, roles and mechanisms in genome stability maintenance].Yi Chuan. 2019 Dec 20;41(12):1084-1098. doi: 10.16288/j.yczz.19-158. Yi Chuan. 2019. PMID: 31857280 Review. Chinese.
Cited by
-
Mechanisms restraining break-induced replication at two-ended DNA double-strand breaks.EMBO J. 2021 May 17;40(10):e104847. doi: 10.15252/embj.2020104847. Epub 2021 Apr 12. EMBO J. 2021. PMID: 33844333 Free PMC article.
-
Loss of Drosophila Mei-41/ATR Alters Meiotic Crossover Patterning.Genetics. 2018 Feb;208(2):579-588. doi: 10.1534/genetics.117.300634. Epub 2017 Dec 15. Genetics. 2018. PMID: 29247012 Free PMC article.
-
DNA Damage Responses during the Cell Cycle: Insights from Model Organisms and Beyond.Genes (Basel). 2021 Nov 25;12(12):1882. doi: 10.3390/genes12121882. Genes (Basel). 2021. PMID: 34946831 Free PMC article. Review.
-
The Drosophila melanogaster Ortholog of RFWD3 Functions Independently of RAD51 During DNA Repair.G3 (Bethesda). 2020 Mar 5;10(3):999-1004. doi: 10.1534/g3.119.400903. G3 (Bethesda). 2020. PMID: 31900333 Free PMC article.
-
A DNA repair protein and histone methyltransferase interact to promote genome stability in the Caenorhabditis elegans germ line.PLoS Genet. 2019 Feb 22;15(2):e1007992. doi: 10.1371/journal.pgen.1007992. eCollection 2019 Feb. PLoS Genet. 2019. PMID: 30794539 Free PMC article.
References
-
- Adams M. D., McVey M., Sekelsky J. J., 2003. Drosophila BLM in double-strand break repair by synthesis-dependent strand annealing. Science 299: 265–267. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
