

Selective expression of TSPAN2 in vascular smooth muscle is independently regulated by TGF-β1/SMAD and myocardin/serum response factor
- PMID: 28258189
- PMCID: PMC5434656
- DOI: 10.1096/fj.201601021R
Selective expression of TSPAN2 in vascular smooth muscle is independently regulated by TGF-β1/SMAD and myocardin/serum response factor
Abstract
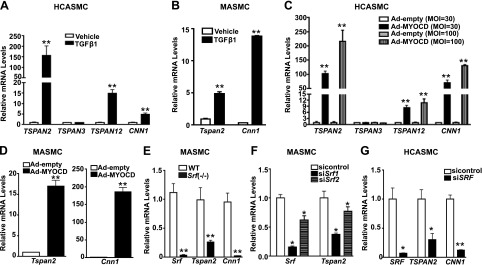
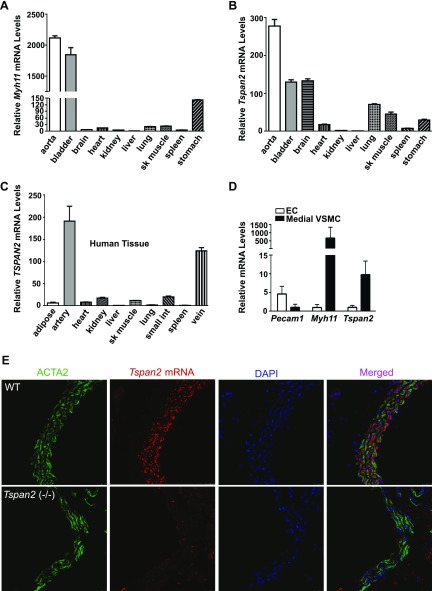
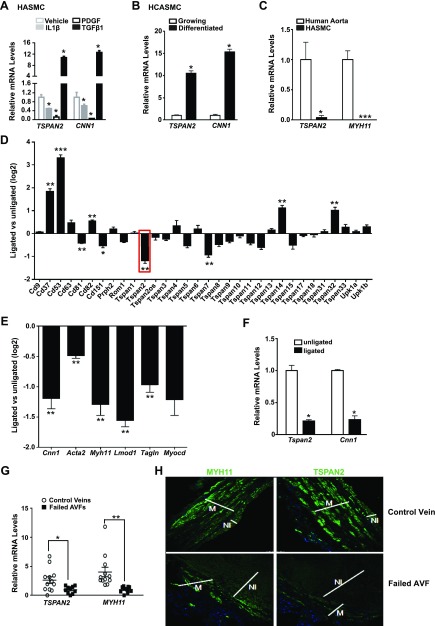
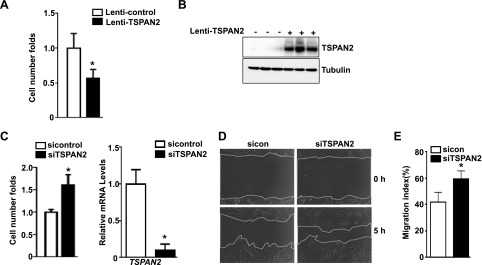
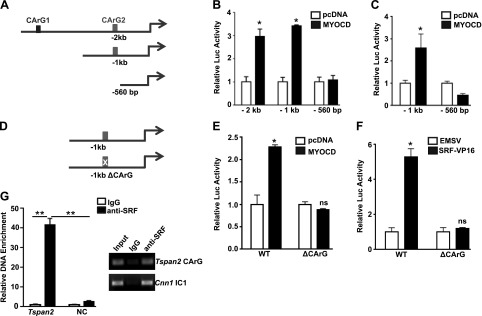
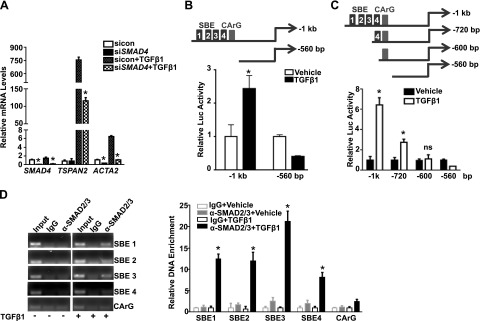
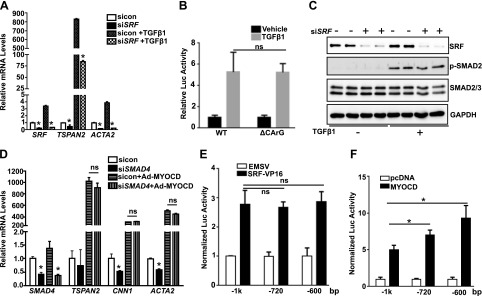
Tetraspanins (TSPANs) comprise a large family of 4-transmembrane domain proteins. The importance of TSPANs in vascular smooth muscle cells (VSMCs) is unexplored. Given that TGF-β1 and myocardin (MYOCD) are potent activators for VSMC differentiation, we screened for TGF-β1 and MYOCD/serum response factor (SRF)-regulated TSPANs in VSMC by using RNA-seq analyses and RNA-arrays. TSPAN2 was found to be the only TSPAN family gene induced by TGF-β1 and MYOCD, and reduced by SRF deficiency in VSMCs. We also found that TSPAN2 is highly expressed in smooth muscle-enriched tissues and down-regulated in in vitro models of VSMC phenotypic modulation. TSPAN2 expression is attenuated in mouse carotid arteries after ligation injury and in failed human arteriovenous fistula samples after occlusion by dedifferentiated neointimal VSMC. In vitro functional studies showed that TSPAN2 suppresses VSMC proliferation and migration. Luciferase reporter and chromatin immunoprecipitation assays demonstrated that TSPAN2 is regulated by 2 parallel pathways, MYOCD/SRF and TGF-β1/SMAD, via distinct binding elements within the proximal promoter. Thus, we identified the first VSMC-enriched and MYOCD/SRF and TGF-β1/SMAD-dependent TSPAN family member, whose expression is intimately associated with VSMC differentiation and negatively correlated with vascular disease. Our results suggest that TSPAN2 may play important roles in vascular disease.-Zhao, J., Wu, W., Zhang, W., Lu, Y. W., Tou, E., Ye, J., Gao, P., Jourd'heuil, D., Singer, H. A., Wu, M., Long, X. Selective expression of TSPAN2 in vascular smooth muscle is independently regulated by TGF-β1/SMAD and myocardin/serum response factor.
Keywords: arteriovenous fistula; differentiation; phenotypic modulation; tetraspanins.
© FASEB.
Figures

Similar articles
-
MYOSLID Is a Novel Serum Response Factor-Dependent Long Noncoding RNA That Amplifies the Vascular Smooth Muscle Differentiation Program.Arterioscler Thromb Vasc Biol. 2016 Oct;36(10):2088-99. doi: 10.1161/ATVBAHA.116.307879. Epub 2016 Jul 21. Arterioscler Thromb Vasc Biol. 2016. PMID: 27444199 Free PMC article.
-
c-Myb Regulates Proliferation and Differentiation of Adventitial Sca1+ Vascular Smooth Muscle Cell Progenitors by Transactivation of Myocardin.Arterioscler Thromb Vasc Biol. 2016 Jul;36(7):1367-76. doi: 10.1161/ATVBAHA.115.307116. Epub 2016 May 12. Arterioscler Thromb Vasc Biol. 2016. PMID: 27174098
-
Enhancer of polycomb1 lessens neointima formation by potentiation of myocardin-induced smooth muscle differentiation.Atherosclerosis. 2012 May;222(1):84-91. doi: 10.1016/j.atherosclerosis.2012.02.015. Epub 2012 Feb 18. Atherosclerosis. 2012. PMID: 22398275
-
Myocardin: A novel player in atherosclerosis.Atherosclerosis. 2017 Feb;257:266-278. doi: 10.1016/j.atherosclerosis.2016.12.002. Epub 2016 Dec 1. Atherosclerosis. 2017. PMID: 28012646 Review.
-
Myocardin and smooth muscle differentiation.Arch Biochem Biophys. 2014 Feb 1;543:48-56. doi: 10.1016/j.abb.2013.12.015. Epub 2013 Dec 25. Arch Biochem Biophys. 2014. PMID: 24374035 Review.
Cited by
-
A Bioinformatics Perspective on the Links Between Tetraspanin-Enriched Microdomains and Cardiovascular Pathophysiology.Front Cardiovasc Med. 2021 Mar 30;8:630471. doi: 10.3389/fcvm.2021.630471. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 33860000 Free PMC article.
-
Transcriptional control of a novel long noncoding RNA Mymsl in smooth muscle cells by a single Cis-element and its initial functional characterization in vessels.J Mol Cell Cardiol. 2020 Jan;138:147-157. doi: 10.1016/j.yjmcc.2019.11.148. Epub 2019 Nov 18. J Mol Cell Cardiol. 2020. PMID: 31751568 Free PMC article.
-
Migrasomes: key players in immune regulation and promising medical applications.Front Immunol. 2025 May 15;16:1592314. doi: 10.3389/fimmu.2025.1592314. eCollection 2025. Front Immunol. 2025. PMID: 40443653 Free PMC article. Review.
-
DNA Methylation and Blood Pressure Phenotypes: A Review of the Literature.Am J Hypertens. 2021 Apr 2;34(3):267-273. doi: 10.1093/ajh/hpab026. Am J Hypertens. 2021. PMID: 33821945 Free PMC article. Review.
-
Unveiling the Potential of Migrasomes: A Machine-Learning-Driven Signature for Diagnosing Acute Myocardial Infarction.Biomedicines. 2024 Jul 22;12(7):1626. doi: 10.3390/biomedicines12071626. Biomedicines. 2024. PMID: 39062199 Free PMC article.
References
-
- Thyberg J. (1998) Phenotypic modulation of smooth muscle cells during formation of neointimal thickenings following vascular injury. Histol. Histopathol. 13, 871–891 - PubMed
-
- Owens G. K., Kumar M. S., Wamhoff B. R. (2004) Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 84, 767–801 - PubMed
-
- Pauletto P., Sarzani R., Rappelli A., Chiavegato A., Pessina A. C., Sartore S. (1994) Differentiation and growth of vascular smooth muscle cells in experimental hypertension. Am. J. Hypertens. 7, 661–674 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
