

Antiviral Resistance Protein Tm-22 Functions on the Plasma Membrane
- PMID: 28258211
- PMCID: PMC5373048
- DOI: 10.1104/pp.16.01512
Antiviral Resistance Protein Tm-22 Functions on the Plasma Membrane
Abstract
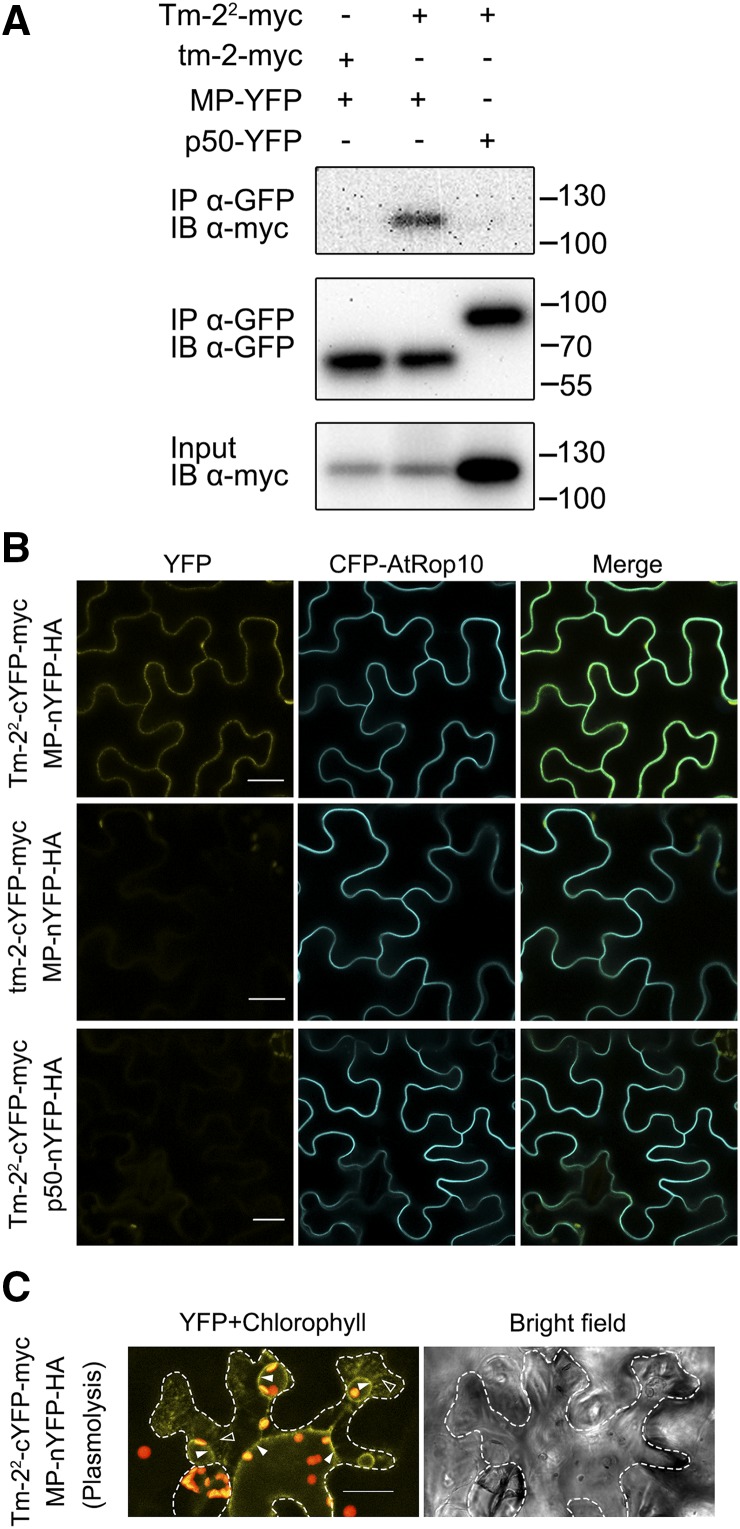
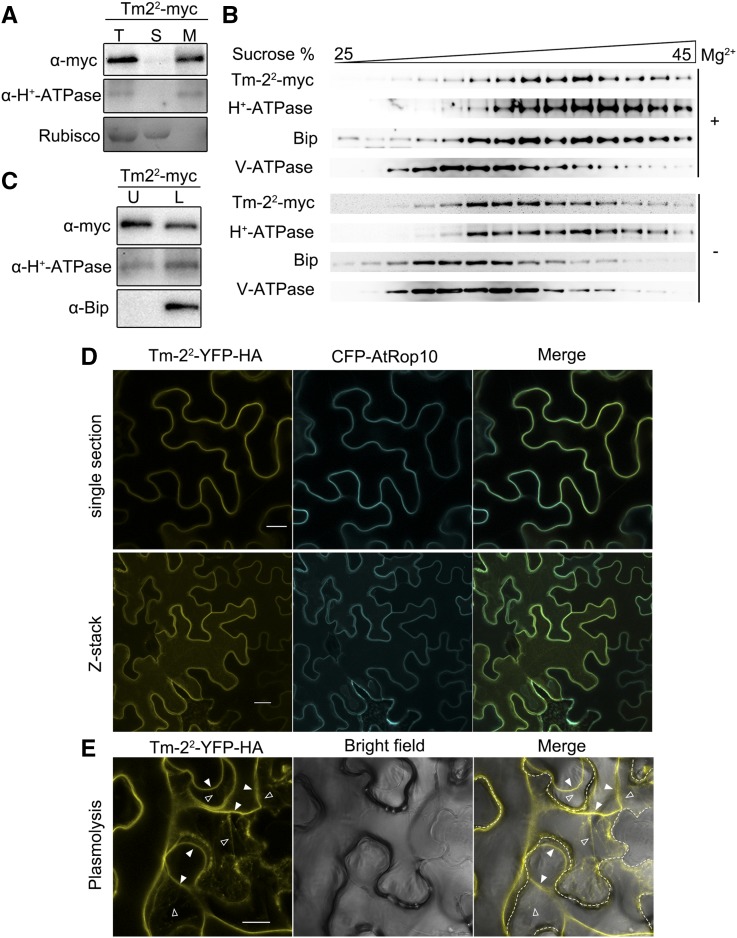
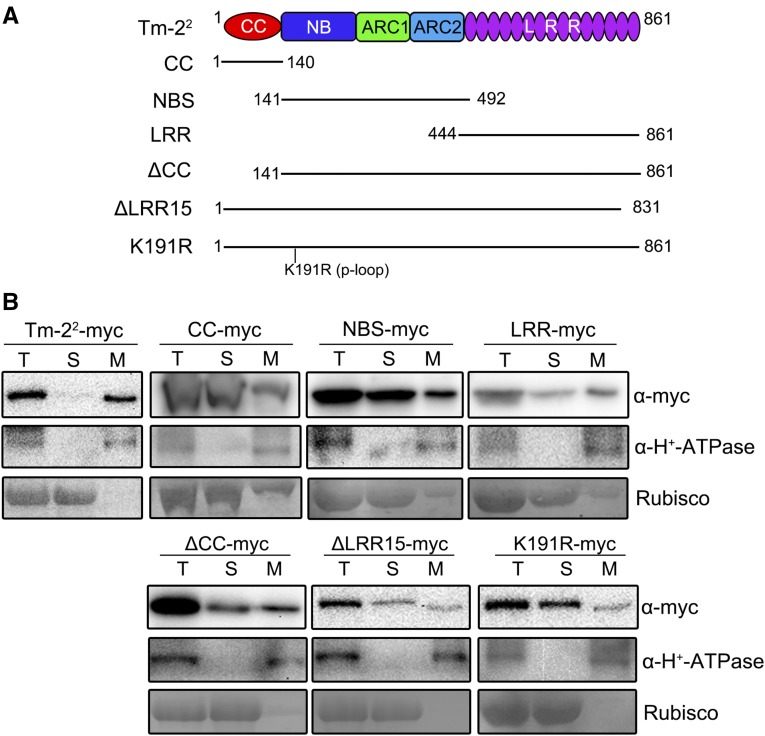
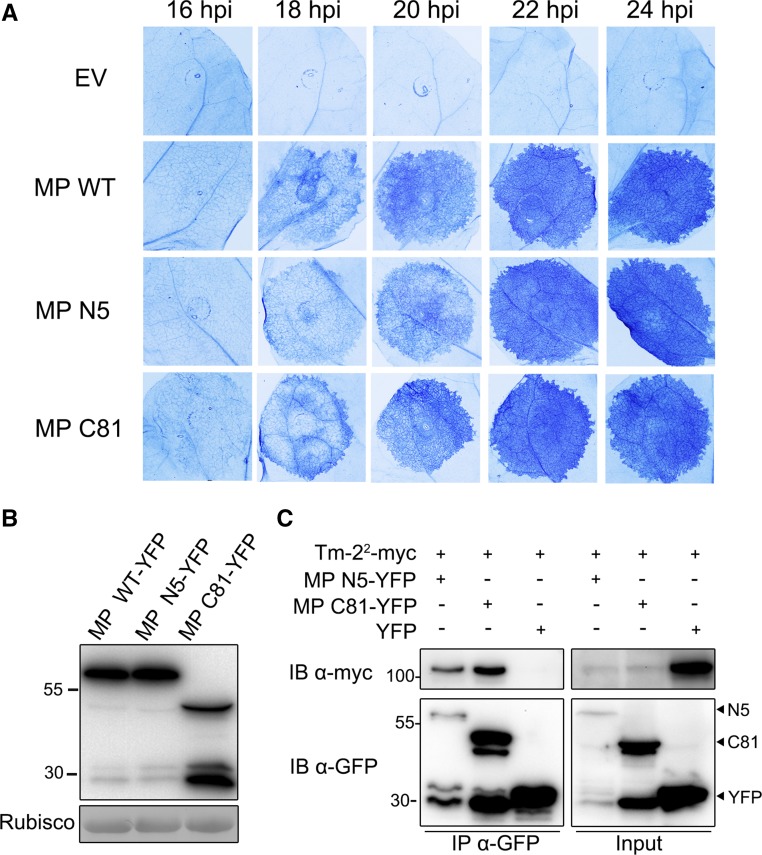
The tomato Tobacco mosaic virus resistance-22 (Tm-22 ) gene encodes a coiled-coil-nucleotide binding site-Leu-rich repeat protein lacking a conventional plasma membrane (PM) localization motif. Tm-22 confers plant extreme resistance against tobamoviruses including Tobacco mosaic virus (TMV) by recognizing the avirulence (Avr) viral movement protein (MP). However, the subcellular compartment where Tm-22 functions is unclear. Here, we demonstrate that Tm-22 interacts with TMV MP to form a protein complex at the PM We show that both inactive and active Tm-22 proteins are localized to the PM When restricted to PM by fusing Tm-22 to the S-acylated PM association motif, the Tm-22 fusion protein can still induce a hypersensitive response cell death, consistent with its activation at the PM Through analyses of viral MP mutants, we find that the plasmodesmata (PD) localization of the Avr protein MP is not required for Tm-22 function. These results suggest that Tm-22-mediated resistance takes place on PM without requirement of its Avr protein to be located to PD.
© 2017 American Society of Plant Biologists. All Rights Reserved.
Figures



References
-
- Axtell MJ, Staskawicz BJ (2003) Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 112: 369–377 - PubMed
-
- Bai S, Liu J, Chang C, Zhang L, Maekawa T, Wang Q, Xiao W, Liu Y, Chai J, Takken FL, Schulze-Lefert P, Shen QH (2012) Structure-function analysis of barley NLR immune receptor MLA10 reveals its cell compartment specific activity in cell death and disease resistance. PLoS Pathog 8: e1002752. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
