

Nicotinic acetylcholine receptors at the single-channel level
- PMID: 28261794
- PMCID: PMC5979820
- DOI: 10.1111/bph.13770
Nicotinic acetylcholine receptors at the single-channel level
Abstract
Over the past four decades, the patch clamp technique and nicotinic ACh (nACh) receptors have established an enduring partnership. Like all good partnerships, each partner has proven significant in its own right, while their union has spurred innumerable advances in life science research. A member and prototype of the superfamily of pentameric ligand-gated ion channels, the nACh receptor is a chemo-electric transducer, binding ACh released from nerves and rapidly opening its channel to cation flow to elicit cellular excitation. A subject of a Nobel Prize in Physiology or Medicine, the patch clamp technique provides unprecedented resolution of currents through single ion channels in their native cellular environments. Here, focusing on muscle and α7 nACh receptors, we describe the extraordinary contribution of the patch clamp technique towards understanding how they activate in response to neurotransmitter, how subtle structural and mechanistic differences among nACh receptor subtypes translate into significant physiological differences, and how nACh receptors are being exploited as therapeutic drug targets.
Linked articles: This article is part of a themed section on Nicotinic Acetylcholine Receptors. To view the other articles in this section visit http://onlinelibrary.wiley.com/doi/10.1111/bph.v175.11/issuetoc/.
© 2017 The British Pharmacological Society.
Figures
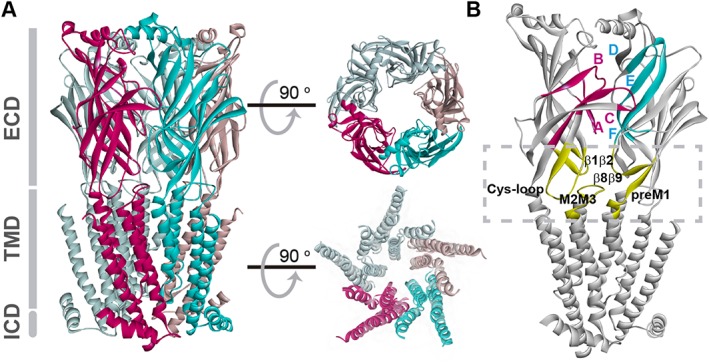
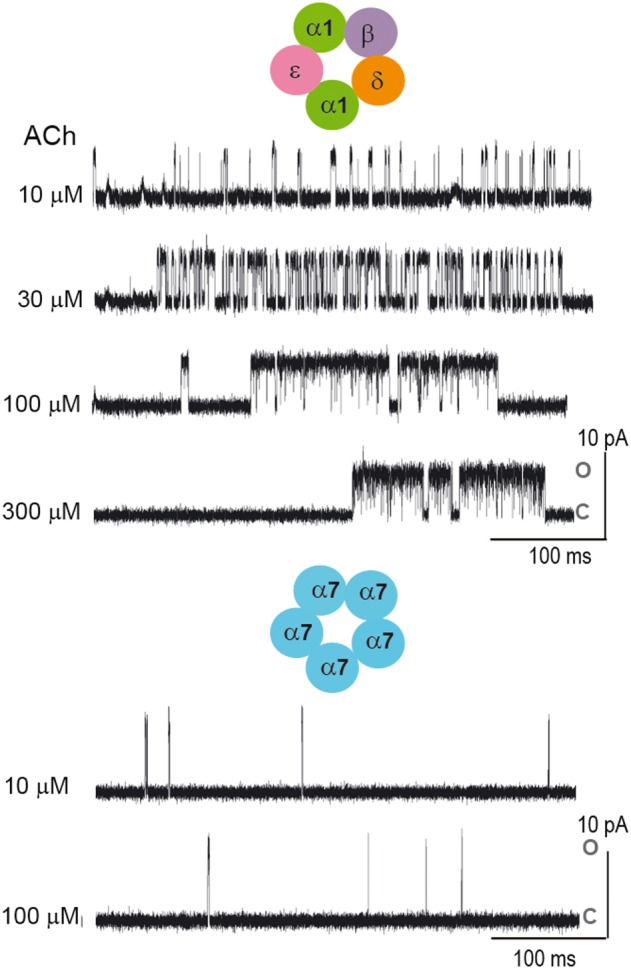
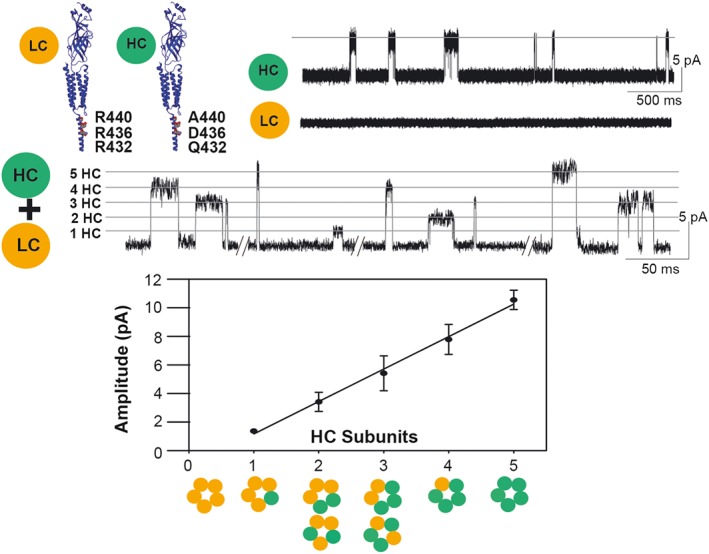
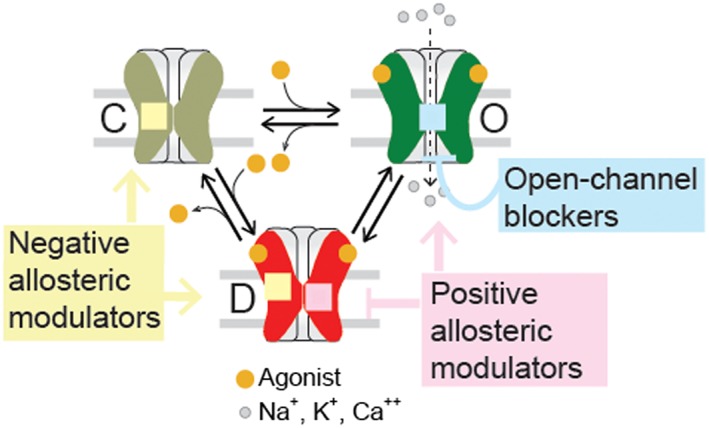
References
-
- Amador M, Dani JA (1991). MK‐801 inhibition of nicotinic acetylcholine receptor channels. Synapse 7: 207–215. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
