

Detecting changes in dynamic and complex acoustic environments
- PMID: 28262095
- PMCID: PMC5367897
- DOI: 10.7554/eLife.24910
Detecting changes in dynamic and complex acoustic environments
Abstract
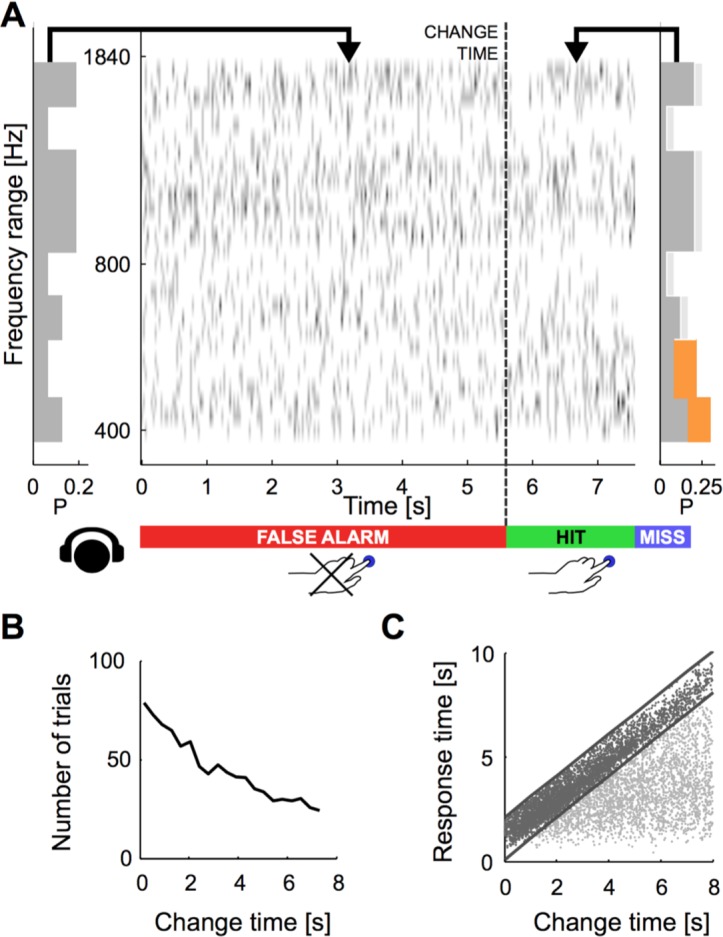
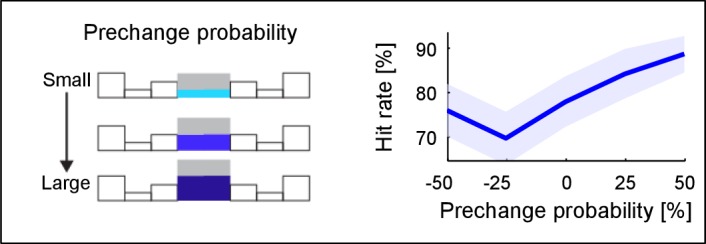
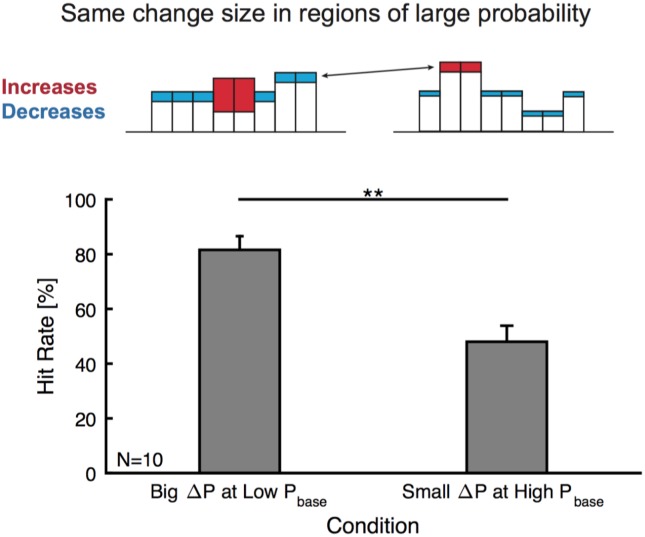
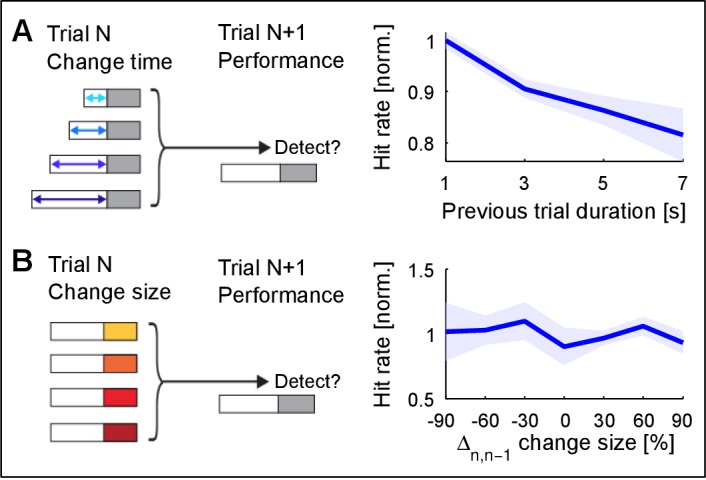
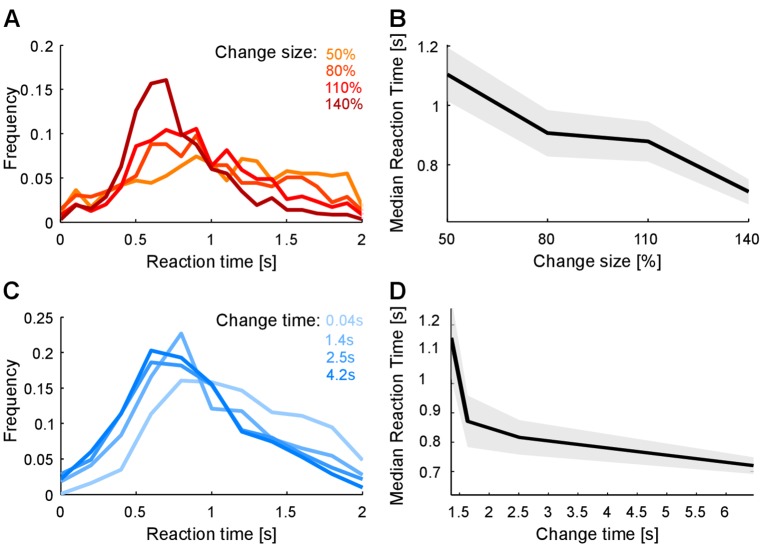
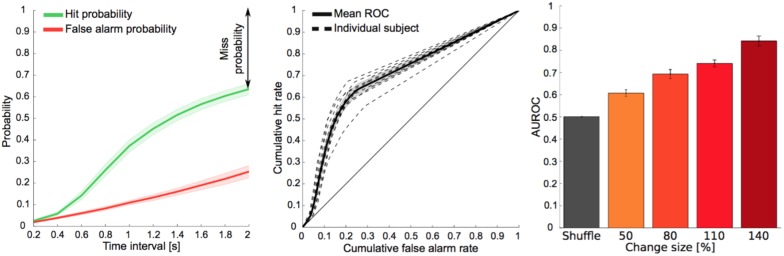
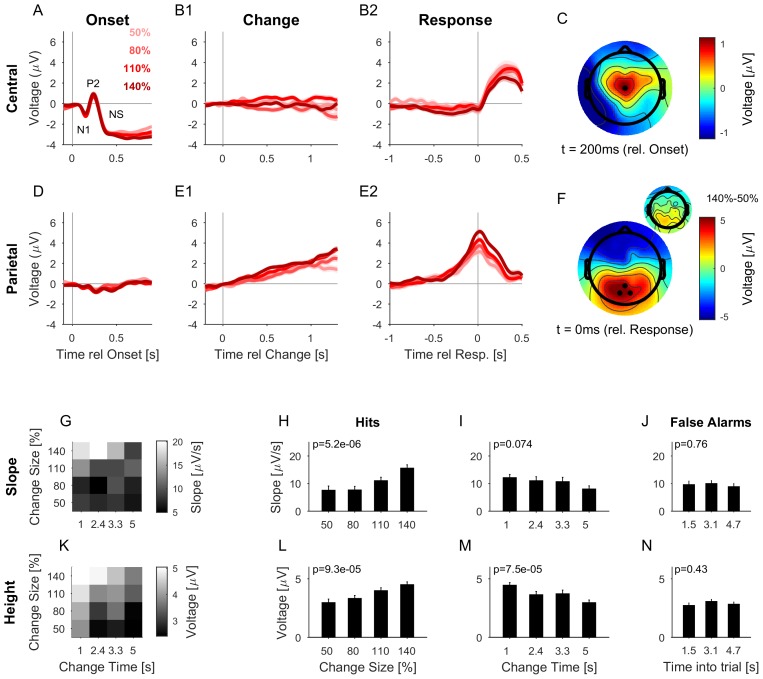
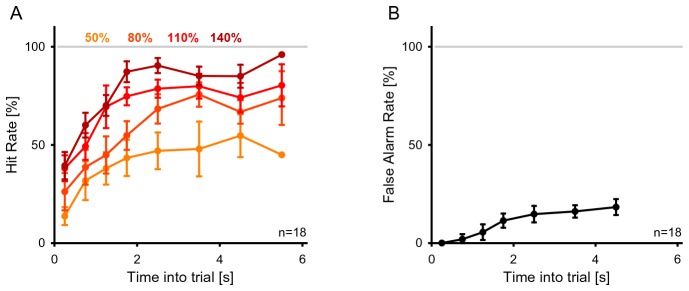
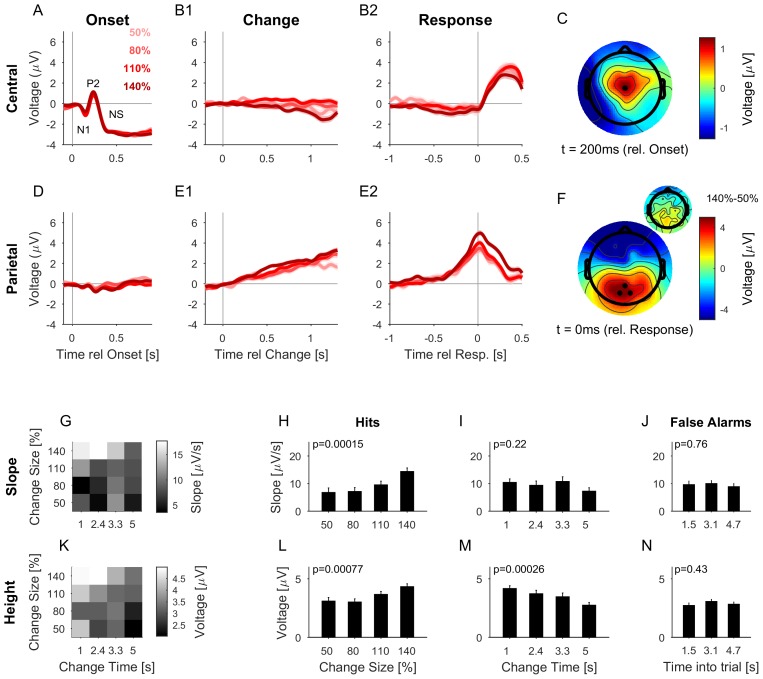
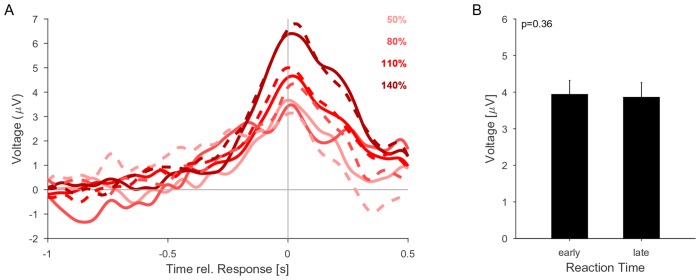
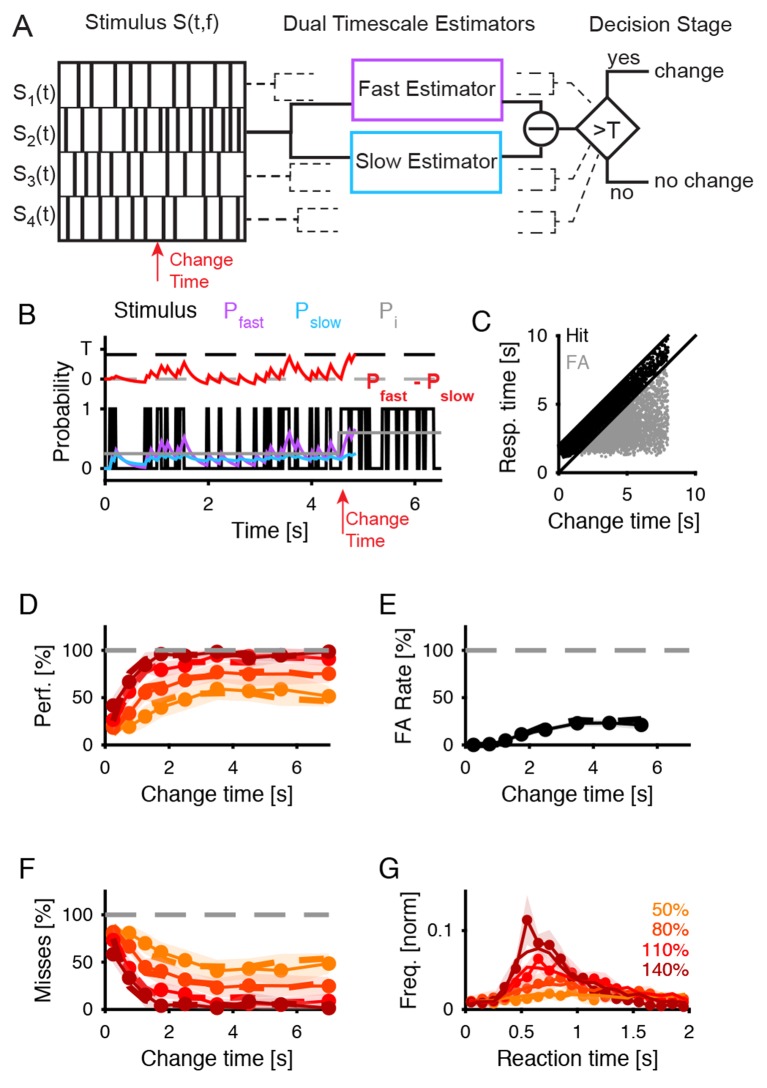
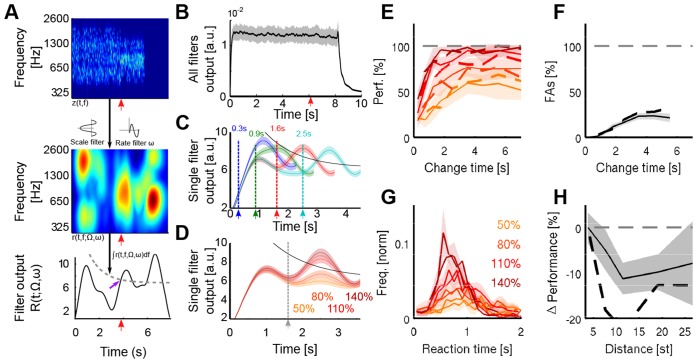
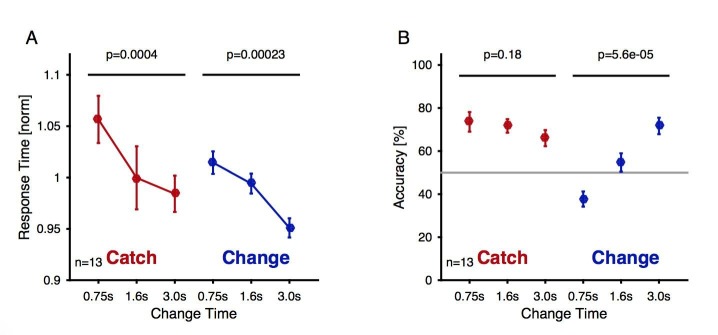
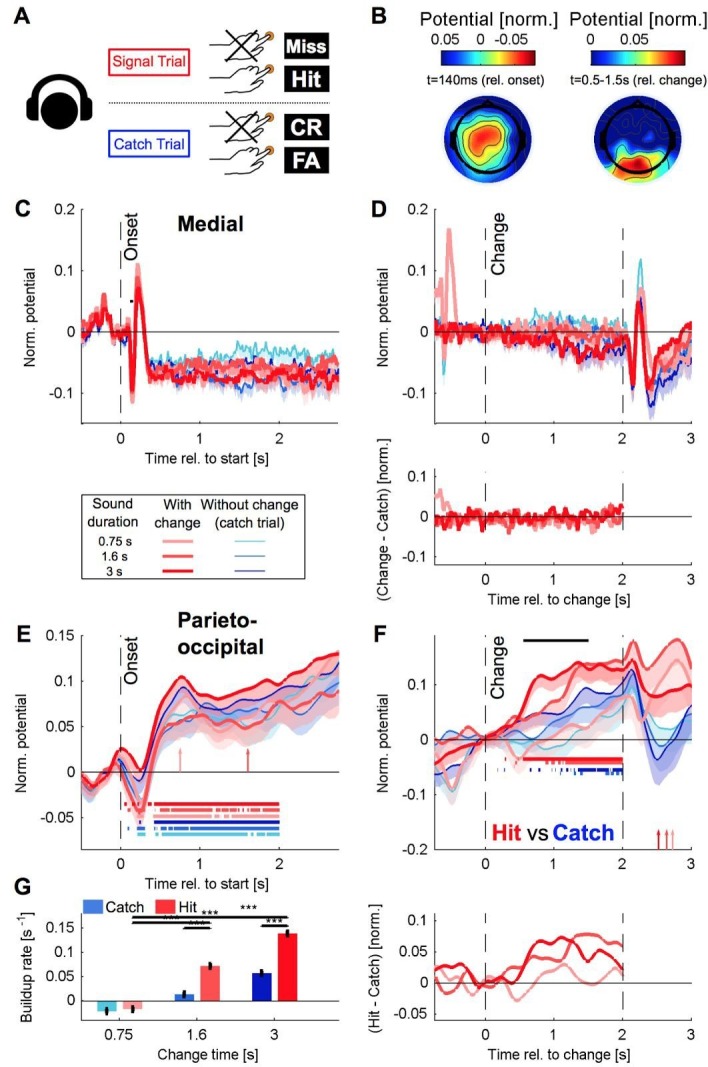
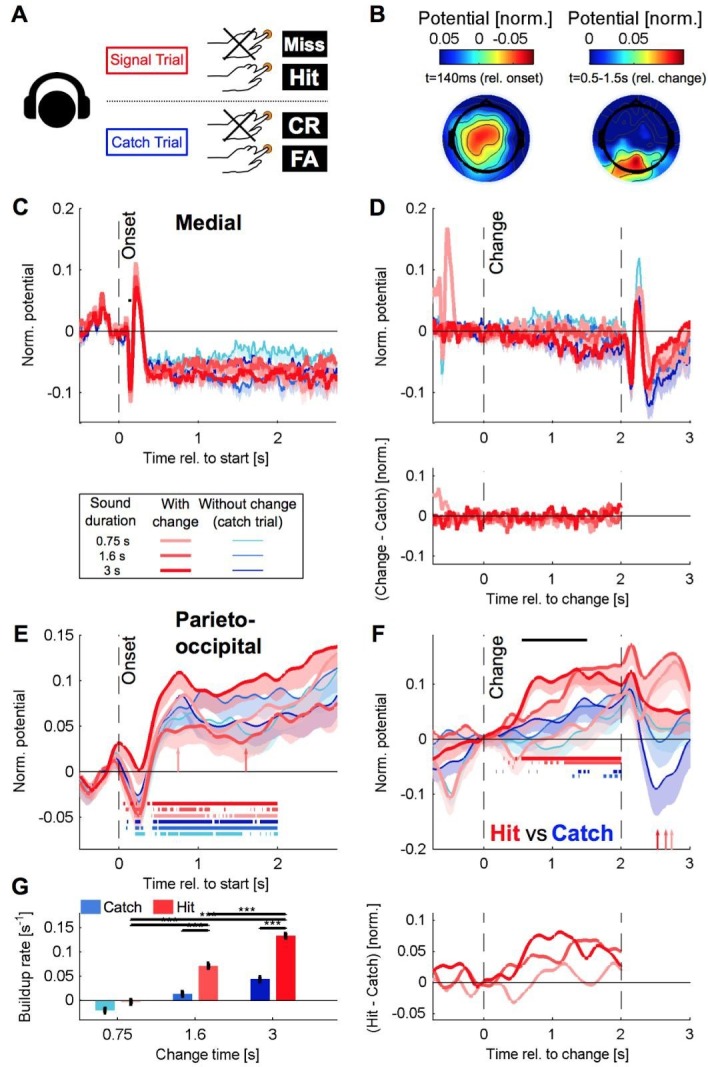
Natural sounds such as wind or rain, are characterized by the statistical occurrence of their constituents. Despite their complexity, listeners readily detect changes in these contexts. We here address the neural basis of statistical decision-making using a combination of psychophysics, EEG and modelling. In a texture-based, change-detection paradigm, human performance and reaction times improved with longer pre-change exposure, consistent with improved estimation of baseline statistics. Change-locked and decision-related EEG responses were found in a centro-parietal scalp location, whose slope depended on change size, consistent with sensory evidence accumulation. The potential's amplitude scaled with the duration of pre-change exposure, suggesting a time-dependent decision threshold. Auditory cortex-related potentials showed no response to the change. A dual timescale, statistical estimation model accounted for subjects' performance. Furthermore, a decision-augmented auditory cortex model accounted for performance and reaction times, suggesting that the primary cortical representation requires little post-processing to enable change-detection in complex acoustic environments.
Keywords: EEG; auditory decision-making; auditory textures; centro-parietal positivity; change detection; human; neuroscience; psychophysics.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures


Similar articles
-
Evidence Integration in Natural Acoustic Textures during Active and Passive Listening.eNeuro. 2018 Apr 13;5(2):ENEURO.0090-18.2018. doi: 10.1523/ENEURO.0090-18.2018. eCollection 2018 Mar-Apr. eNeuro. 2018. PMID: 29662943 Free PMC article.
-
Neural Encoding of Auditory Statistics.J Neurosci. 2021 Aug 4;41(31):6726-6739. doi: 10.1523/JNEUROSCI.1887-20.2021. Epub 2021 Jun 30. J Neurosci. 2021. PMID: 34193552 Free PMC article.
-
Change Detection in Auditory Textures.Adv Exp Med Biol. 2016;894:229-239. doi: 10.1007/978-3-319-25474-6_24. Adv Exp Med Biol. 2016. PMID: 27080663 Free PMC article.
-
Functional imaging of auditory scene analysis.Hear Res. 2014 Jan;307:98-110. doi: 10.1016/j.heares.2013.08.003. Epub 2013 Aug 19. Hear Res. 2014. PMID: 23968821 Review.
-
How do auditory cortex neurons represent communication sounds?Hear Res. 2013 Nov;305:102-12. doi: 10.1016/j.heares.2013.03.011. Epub 2013 Apr 17. Hear Res. 2013. PMID: 23603138 Review.
Cited by
-
Pupillary dynamics reflect the impact of temporal expectation on detection strategy.iScience. 2023 Jan 16;26(2):106000. doi: 10.1016/j.isci.2023.106000. eCollection 2023 Feb 17. iScience. 2023. PMID: 36798438 Free PMC article.
-
Signal integration at spherical bushy cells enhances representation of temporal structure but limits its range.Elife. 2017 Sep 25;6:e29639. doi: 10.7554/eLife.29639. Elife. 2017. PMID: 28945194 Free PMC article.
-
Neural correlates of perceptual texture change during active touch.Front Neurosci. 2023 Jun 2;17:1197113. doi: 10.3389/fnins.2023.1197113. eCollection 2023. Front Neurosci. 2023. PMID: 37332863 Free PMC article.
-
Computational framework for investigating predictive processing in auditory perception.J Neurosci Methods. 2021 Aug 1;360:109177. doi: 10.1016/j.jneumeth.2021.109177. Epub 2021 Apr 9. J Neurosci Methods. 2021. PMID: 33839191 Free PMC article.
-
Illusory sound texture reveals multi-second statistical completion in auditory scene analysis.Nat Commun. 2019 Nov 8;10(1):5096. doi: 10.1038/s41467-019-12893-0. Nat Commun. 2019. PMID: 31704913 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
