

Inefficient Crossover Maturation Underlies Elevated Aneuploidy in Human Female Meiosis
- PMID: 28262352
- PMCID: PMC5408880
- DOI: 10.1016/j.cell.2017.02.002
Inefficient Crossover Maturation Underlies Elevated Aneuploidy in Human Female Meiosis
Abstract
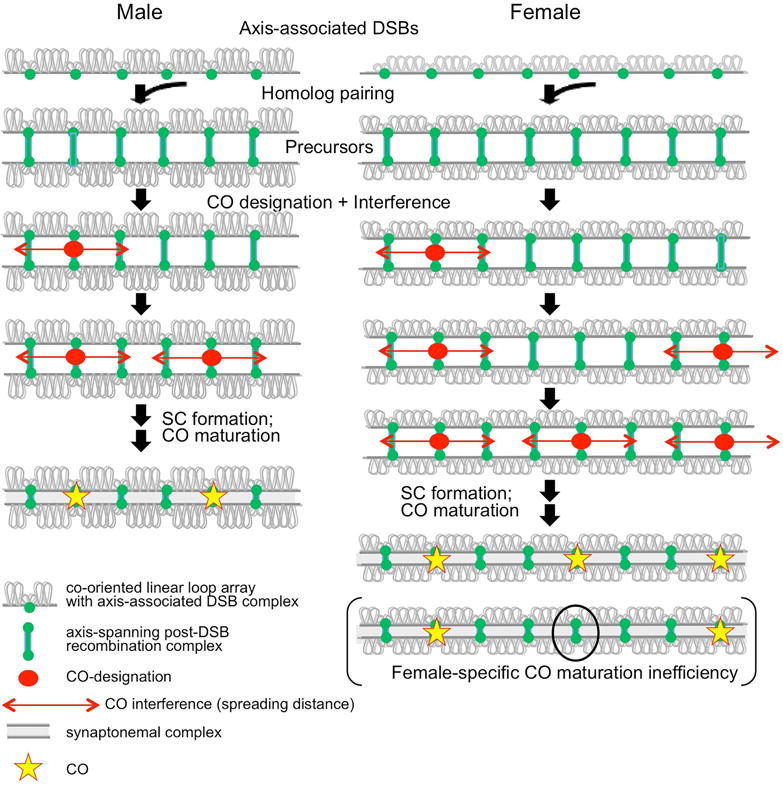
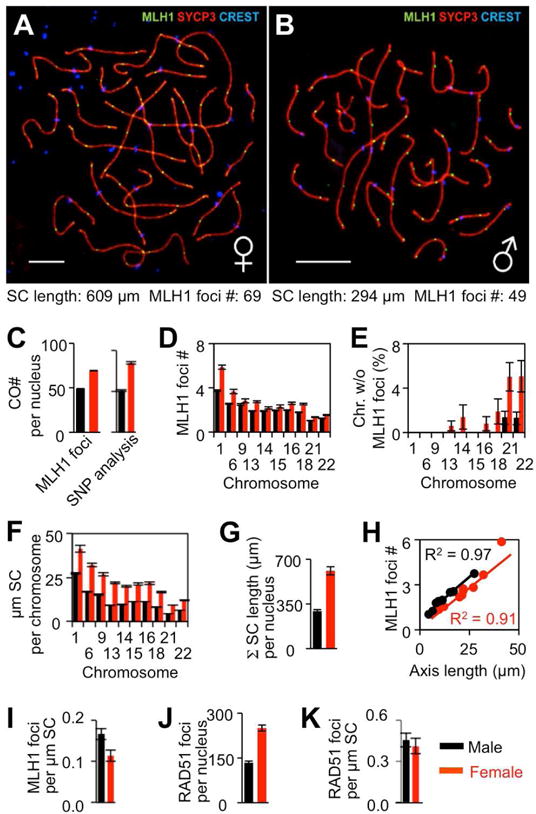
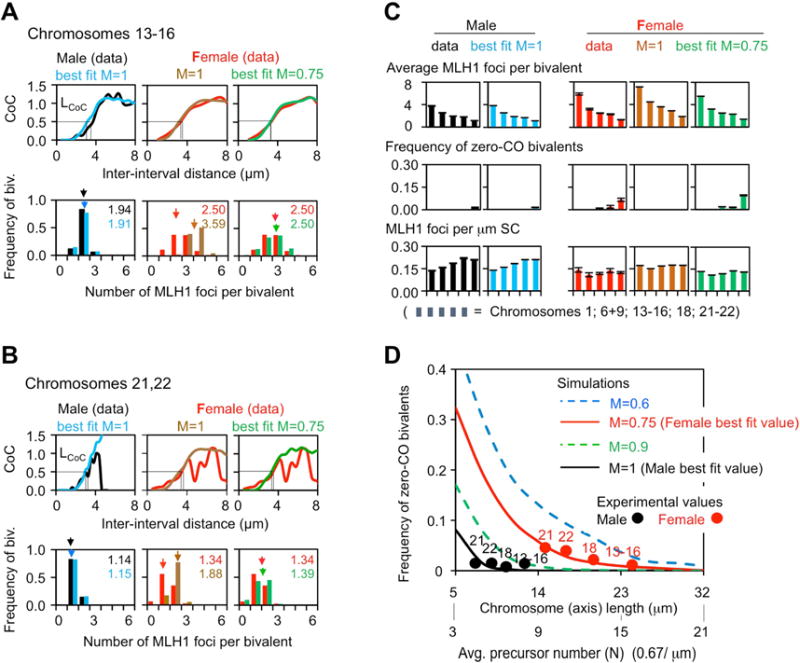
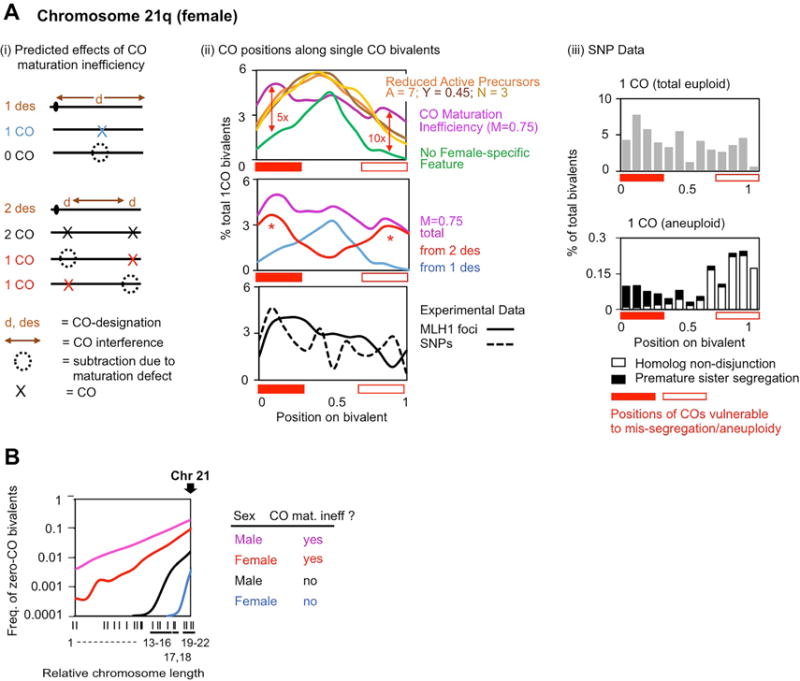
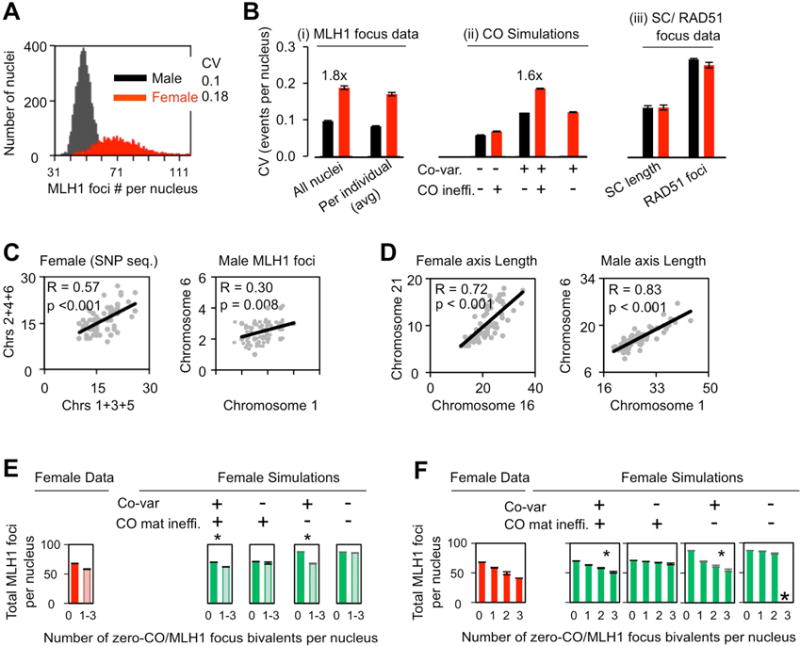
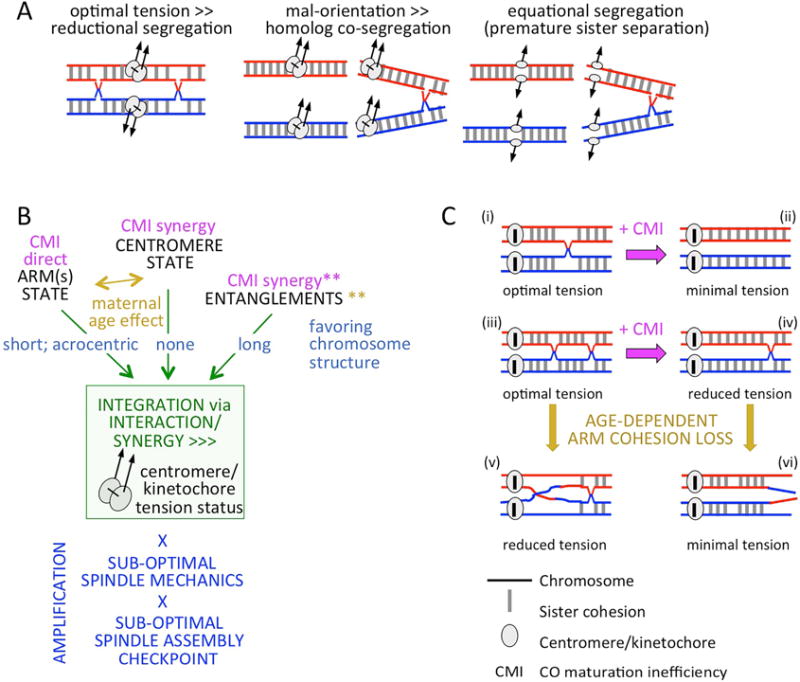
Meiosis is the cellular program that underlies gamete formation. For this program, crossovers between homologous chromosomes play an essential mechanical role to ensure regular segregation. We present a detailed study of crossover formation in human male and female meiosis, enabled by modeling analysis. Results suggest that recombination in the two sexes proceeds analogously and efficiently through most stages. However, specifically in female (but not male), ∼25% of the intermediates that should mature into crossover products actually fail to do so. Further, this "female-specific crossover maturation inefficiency" is inferred to make major contributions to the high level of chromosome mis-segregation and resultant aneuploidy that uniquely afflicts human female oocytes (e.g., giving Down syndrome). Additionally, crossover levels on different chromosomes in the same nucleus tend to co-vary, an effect attributable to global per-nucleus modulation of chromatin loop size. Maturation inefficiency could potentially reflect an evolutionary advantage of increased aneuploidy for human females.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Bojko B. Human meiosis VIII: chromosome pairing and formation of the synaptonemal complex in oocytes. Carisberg Res Commun. 1983;48:457–483.
-
- Bojko M. Human meiosis IX. Crossing over and chiasma formation in oocytes. Carisberg Res Commun. 1985;50:43–72.
Complete references for Data Sources
-
- Codina-Pascual M, Campillo M, Kraus J, Speicher MR, Egozcue J, Navarro J, Benet J. Crossover frequency and synaptonemal complex length: their variability and effects on human male meiosis. Mol Hum Reprod. 2006;12:123–133. - PubMed
-
- Coop G, Wen X, Ober C, Pritchard JK, Przeworski M. High-resolution mapping of crossovers reveals extensive variation in fine-scale recombination patterns among humans. Science. 2008;319:1395–1398. - PubMed
-
- Hassold T, Judis L, Chan ER, Schwartz S, Seftel A, Lynn A. Cytological studies of meiotic recombination in human males. Cytogenet Genome Res. 2004;107:249–255. - PubMed
- Hou Y, Fan W, Yan L, Li R, Lian Y, Huang J, Li J, Xu L, Tang F, Xie XS, Qiao J. Genome analyses of single human oocytes. Cell. 2013;155:1492–506. - PubMed
-
- Kong A, Barnard J, Gudbjartsson DF, Thorleifsson G, Jonsdottir G, Sigurdardottir S, Richardsson B, Jonsdottir J, Thorgeirsson T, Frigge ML, et al. Recombination rate and reproductive success in humans. Nat Genet. 2004;36:1203–1206. - PubMed
-
- Pan Z, Yang Q, Ye N, Wang L, Li J, Yu D, Cooke HJ, Shi Q. Complex relationship between meiotic recombination frequency and autosomal synaptonemal complex length per cell in normal human males. Am J Med Genet. 2012;158A:581–587. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
