

Collective Infectious Units in Viruses
- PMID: 28262512
- PMCID: PMC5837019
- DOI: 10.1016/j.tim.2017.02.003
Collective Infectious Units in Viruses
Abstract
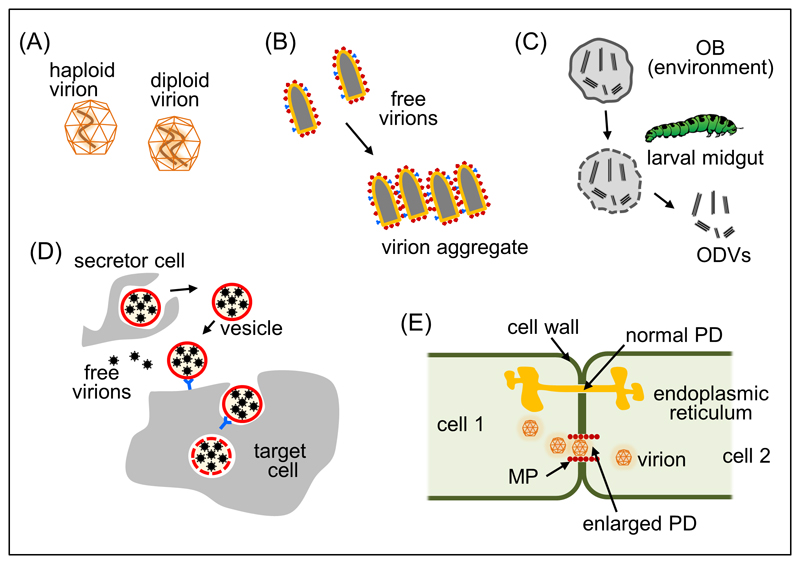
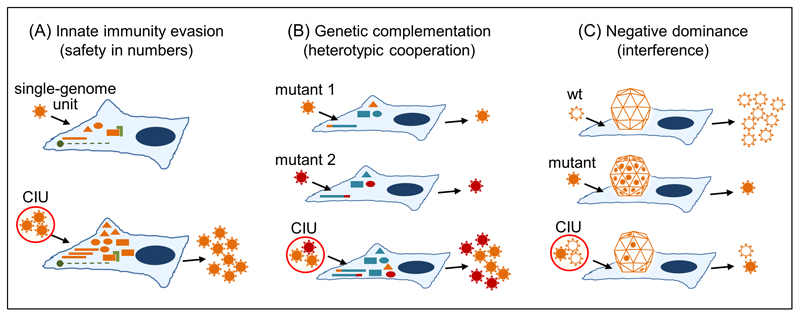
Increasing evidence indicates that viruses do not simply propagate as independent virions among cells, organs, and hosts. Instead, viral spread is often mediated by structures that simultaneously transport groups of viral genomes, such as polyploid virions, aggregates of virions, virion-containing proteinaceous structures, secreted lipid vesicles, and virus-induced cell-cell contacts. These structures increase the multiplicity of infection, independently of viral population density and transmission bottlenecks. Collective infectious units may contribute to the maintenance of viral genetic diversity, and could have implications for the evolution of social-like virus-virus interactions. These may include various forms of cooperation such as immunity evasion, genetic complementation, division of labor, and relaxation of fitness trade-offs, but also noncooperative interactions such as negative dominance and interference, potentially leading to conflict.
Keywords: Baculoviruses; Genetic diversity; Microvesicles; Multiplicity of infection; Polyploid virion; Social evolution.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- López J, Webster RE. Morphogenesis of filamentous bacteriophage f1: orientation of extrusion and production of polyphage. Virology. 1983;127:177–193. - PubMed
-
- Iglesias-Sánchez MJ, López-Galíndez C. Each genomic RNA in HIV-1 heterozygous virus generate new virions. Virology. 2005;333:316–323. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
