

SHH E176/E177-Zn2+ conformation is required for signaling at endogenous sites
- PMID: 28263766
- PMCID: PMC6047533
- DOI: 10.1016/j.ydbio.2017.02.006
SHH E176/E177-Zn2+ conformation is required for signaling at endogenous sites
Abstract
Sonic hedgehog (SHH) is a master developmental regulator. In 1995, the SHH crystal structure predicted that SHH-E176 (human)/E177 (mouse) regulates signaling through a Zn2+-dependent mechanism. While Zn2+ is known to be required for SHH protein stability, a regulatory role for SHH-E176 or Zn2+ has not been described. Here, we show that SHH-E176/177 modulates Zn2+-dependent cross-linking in vitro and is required for endogenous signaling, in vivo. While ectopically expressed SHH-E176A is highly active, mice expressing SHH-E177A at endogenous sites (ShhE177A/-) are morphologically indistinguishable from mice lacking SHH (Shh-/-), with patterning defects in both embryonic spinal cord and forebrain. SHH-E177A distribution along the embryonic spinal cord ventricle is unaltered, suggesting that E177 does not control long-range transport. While SHH-E177A association with cilia basal bodies increases in embryonic ventral spinal cord, diffusely distributed SHH-E177A is not detected. Together, these results reveal a novel role for E177-Zn2+ in regulating SHH signaling that may involve critical, cilia basal-body localized changes in cross-linking and/or conformation.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
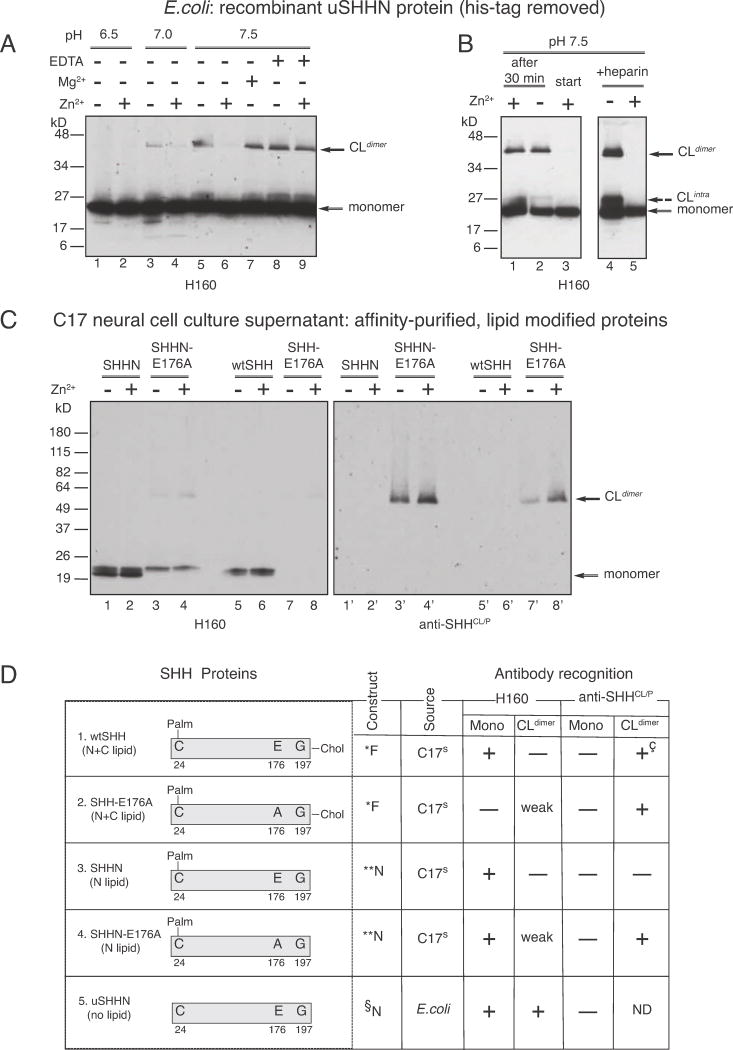
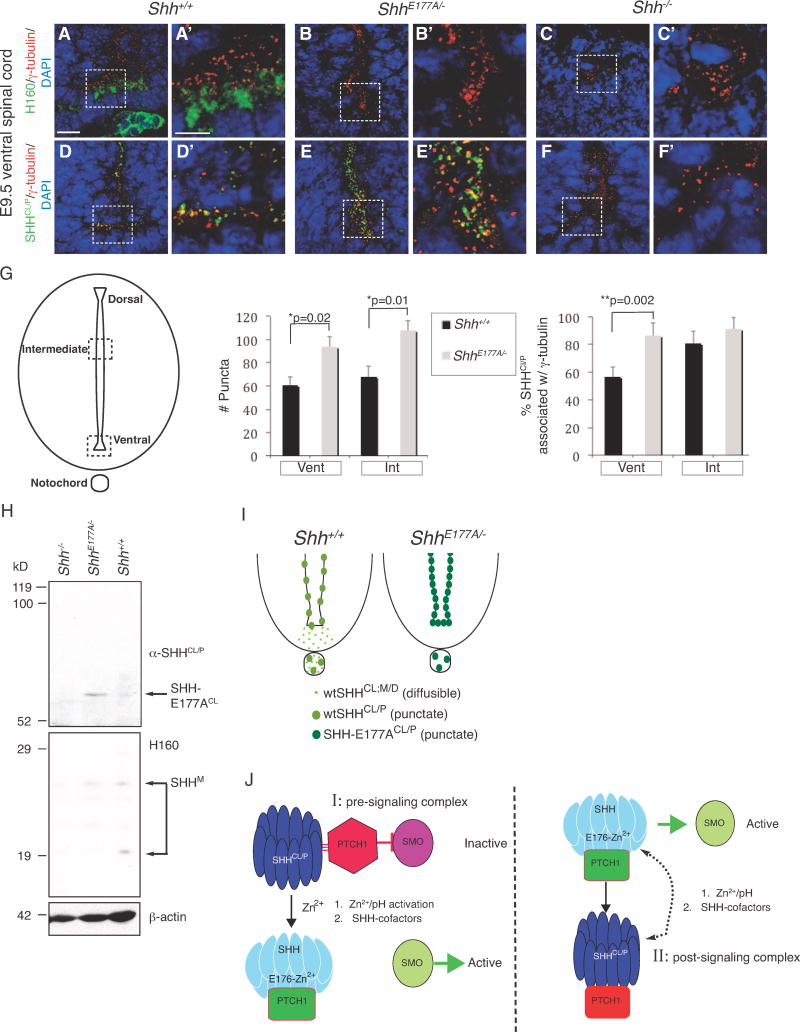
E176A/E177A modulates Zn2+-mediated conformational change, detected by increased formation of cross-linked dimers.
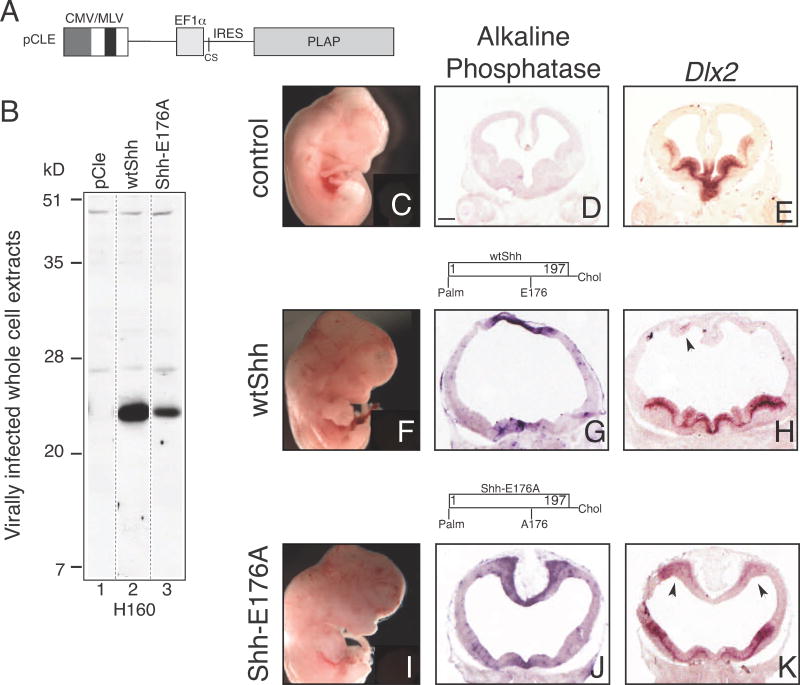
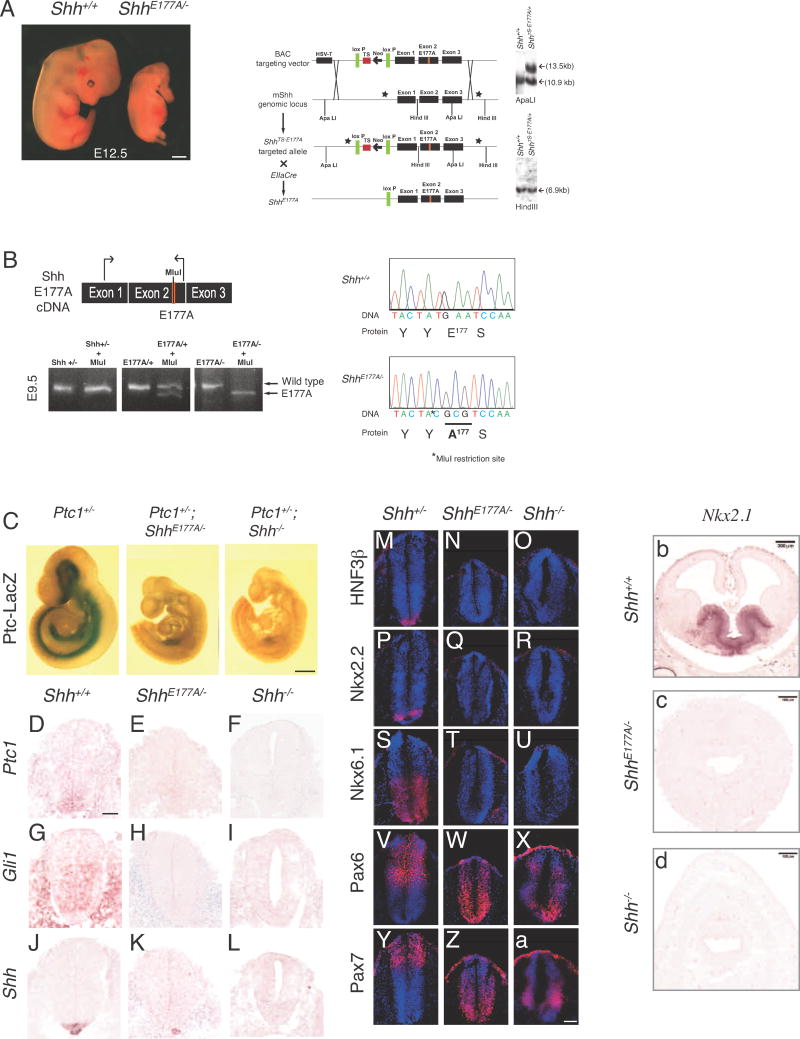
E176A/E177A is active at ectopic, but not endogenous sites in vivo.
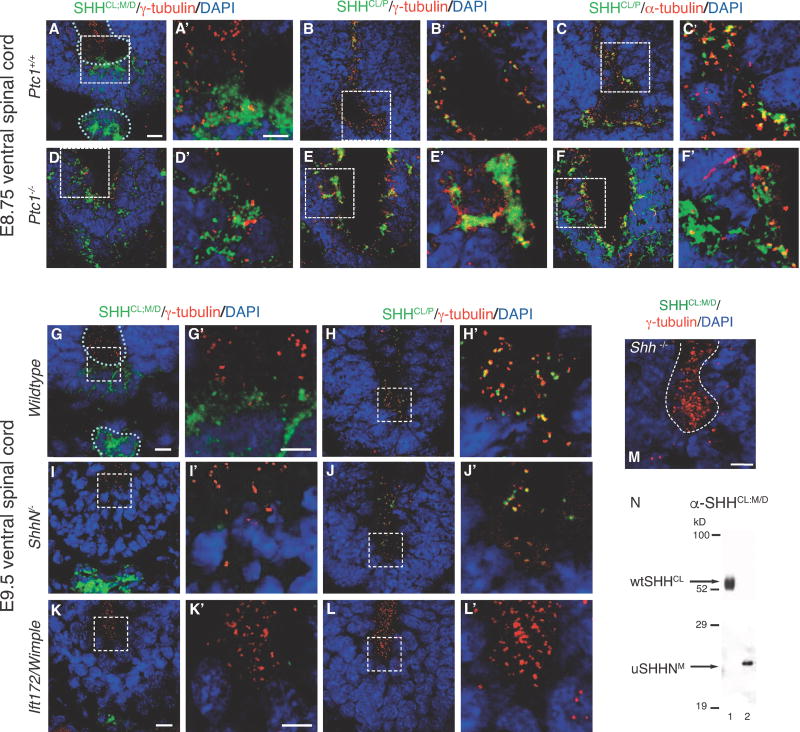
In ShhE177A/− mutant spinal cord, increased accumulation of SHH-E177A occurs near cilia BBs.
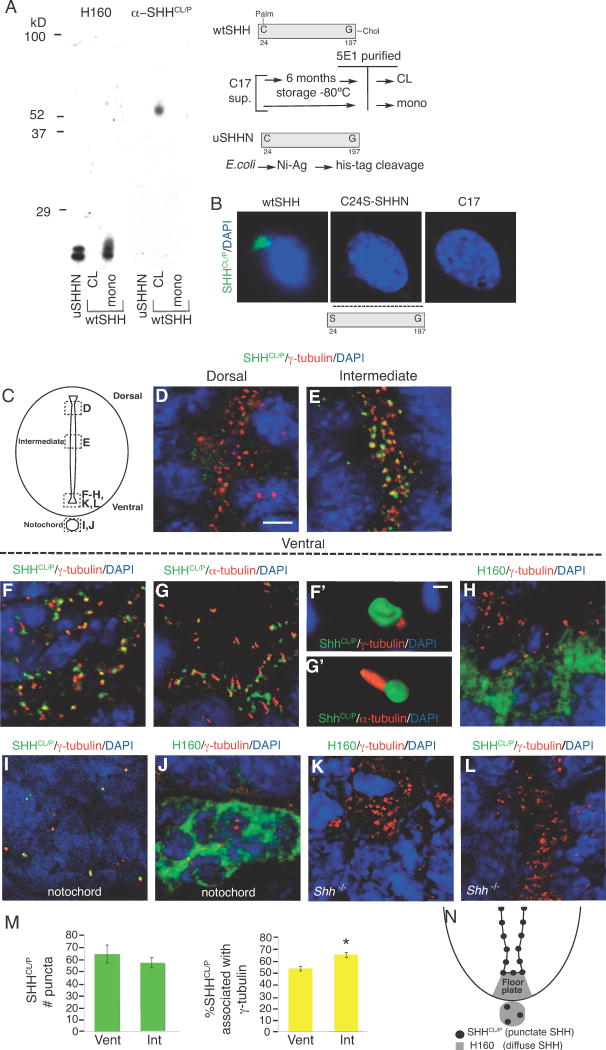
SHHCL/P is still present in the ventral spinal cord even in the absence of the SHH receptor PTC1, supporting a pre-signaling role of SHHCL/P.
Similar articles
-
Tubby-like protein 3 (TULP3) regulates patterning in the mouse embryo through inhibition of Hedgehog signaling.Hum Mol Genet. 2009 May 15;18(10):1740-54. doi: 10.1093/hmg/ddp113. Epub 2009 Mar 12. Hum Mol Genet. 2009. PMID: 19286674 Free PMC article.
-
Spop regulates Gli3 activity and Shh signaling in dorsoventral patterning of the mouse spinal cord.Dev Biol. 2017 Dec 1;432(1):72-85. doi: 10.1016/j.ydbio.2017.04.002. Epub 2017 Apr 12. Dev Biol. 2017. PMID: 28412462 Free PMC article.
-
Primary cilia are critical for Sonic hedgehog-mediated dopaminergic neurogenesis in the embryonic midbrain.Dev Biol. 2016 Jan 1;409(1):55-71. doi: 10.1016/j.ydbio.2015.10.033. Epub 2015 Nov 2. Dev Biol. 2016. PMID: 26542012 Free PMC article.
-
Essential roles of Gli3 and sonic hedgehog in pattern formation and developmental anomalies caused by their dysfunction.Congenit Anom (Kyoto). 2006 Sep;46(3):123-8. doi: 10.1111/j.1741-4520.2006.00114.x. Congenit Anom (Kyoto). 2006. PMID: 16922918 Review.
-
Hedgehog trafficking, cilia and brain functions.Differentiation. 2012 Feb;83(2):S97-104. doi: 10.1016/j.diff.2011.11.011. Epub 2011 Dec 9. Differentiation. 2012. PMID: 22169886 Review.
Cited by
-
Manipulation of Sonic Hedgehog Signaling Pathway in Maintenance, Differentiation, and Endocrine Activity of Insulin-Producing Cells: A Systematic Review.Iran J Med Sci. 2024 Feb 1;49(2):65-76. doi: 10.30476/ijms.2023.95425.2678. eCollection 2024 Feb. Iran J Med Sci. 2024. PMID: 38356490 Free PMC article.
-
Gain-of-function Shh mutants activate Smo cell-autonomously independent of Ptch1/2 function.Mech Dev. 2018 Oct;153:30-41. doi: 10.1016/j.mod.2018.08.009. Epub 2018 Aug 23. Mech Dev. 2018. PMID: 30144507 Free PMC article.
-
Machine learning differentiates enzymatic and non-enzymatic metals in proteins.Nat Commun. 2021 Jun 17;12(1):3712. doi: 10.1038/s41467-021-24070-3. Nat Commun. 2021. PMID: 34140507 Free PMC article.
-
Inactive metallopeptidase homologs: the secret lives of pseudopeptidases.Front Mol Biosci. 2024 Jul 10;11:1436917. doi: 10.3389/fmolb.2024.1436917. eCollection 2024. Front Mol Biosci. 2024. PMID: 39050735 Free PMC article. Review.
-
Association of Sonic Hedgehog with the extracellular matrix requires its zinc-coordination center.BMC Mol Cell Biol. 2021 Apr 16;22(1):22. doi: 10.1186/s12860-021-00359-5. BMC Mol Cell Biol. 2021. PMID: 33863273 Free PMC article.
References
-
- Briscoe J, Ericson J. Specification of neuronal fates in the ventral neural tube. Curr Opin Neurobiol. 2001;11:43–49. - PubMed
-
- Chamberlain CE, Jeong J, Guo C, Allen BL, McMahon AP. Notochord-derived Shh concentrates in close association with the apically positioned basal body in neural target cells and forms a dynamic gradient during neural patterning. Development. 2008;135:1097–1106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
