

Plasma Membrane Na⁺-Coupled Citrate Transporter (SLC13A5) and Neonatal Epileptic Encephalopathy
- PMID: 28264506
- PMCID: PMC6155422
- DOI: 10.3390/molecules22030378
Plasma Membrane Na⁺-Coupled Citrate Transporter (SLC13A5) and Neonatal Epileptic Encephalopathy
Abstract
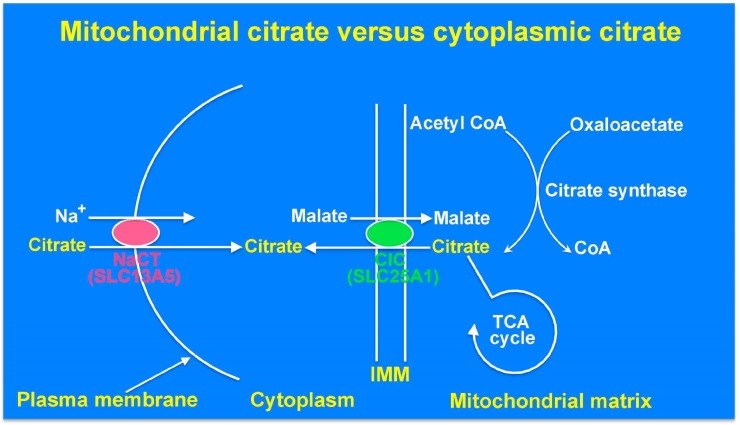
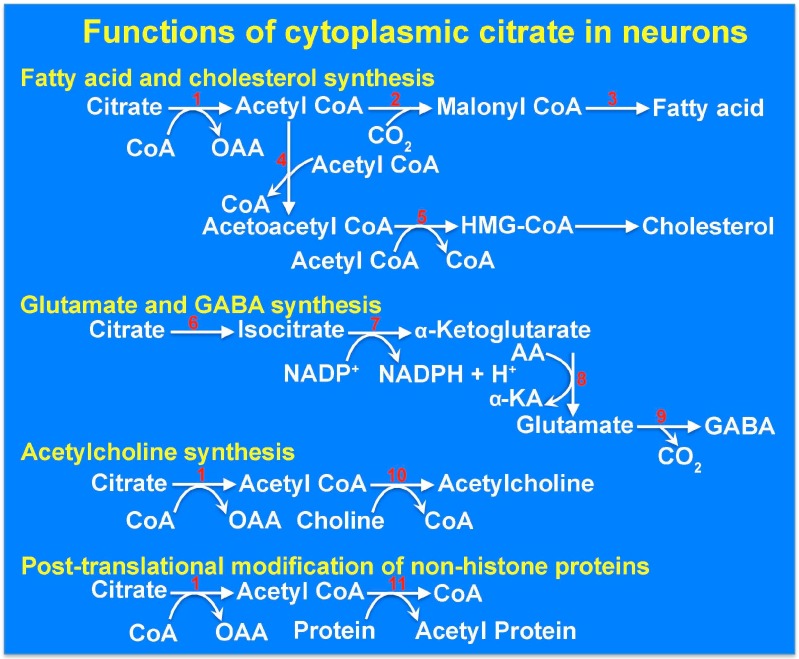
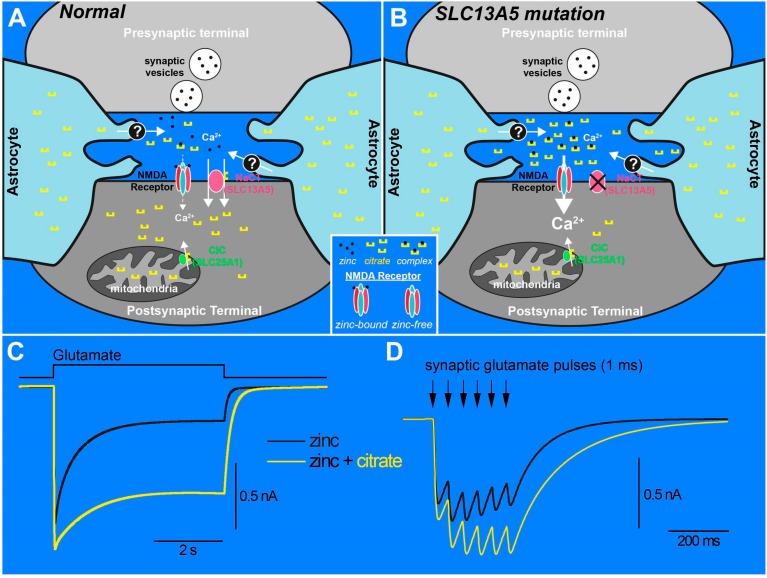
SLC13A5 is a Na⁺-coupled transporter for citrate that is expressed in the plasma membrane of specific cell types in the liver, testis, and brain. It is an electrogenic transporter with a Na⁺:citrate3- stoichiometry of 4:1. In humans, the Michaelis constant for SLC13A5 to transport citrate is ~600 μM, which is physiologically relevant given that the normal concentration of citrate in plasma is in the range of 150-200 μM. Li⁺ stimulates the transport function of human SLC13A5 at concentrations that are in the therapeutic range in patients on lithium therapy. Human SLC13A5 differs from rodent Slc13a5 in two important aspects: the affinity of the human transporter for citrate is ~30-fold less than that of the rodent transporter, thus making human SLC13A5 a low-affinity/high-capacity transporter and the rodent Slc13a5 a high-affinity/low-capacity transporter. In the liver, SLC13A5 is expressed exclusively in the sinusoidal membrane of the hepatocytes, where it plays a role in the uptake of circulating citrate from the sinusoidal blood for metabolic use. In the testis, the transporter is expressed only in spermatozoa, which is also only in the mid piece where mitochondria are located; the likely function of the transporter in spermatozoa is to mediate the uptake of citrate present at high levels in the seminal fluid for subsequent metabolism in the sperm mitochondria to generate biological energy, thereby supporting sperm motility. In the brain, the transporter is expressed mostly in neurons. As astrocytes secrete citrate into extracellular medium, the potential function of SLC13A5 in neurons is to mediate the uptake of circulating citrate and astrocyte-released citrate for subsequent metabolism. Slc13a5-knockout mice have been generated; these mice do not have any overt phenotype but are resistant to experimentally induced metabolic syndrome. Recently however, loss-of-function mutations in human SLC13A5 have been found to cause severe epilepsy and encephalopathy early in life. Interestingly, there is no evidence of epilepsy or encephalopathy in Slc13a5-knockout mice, underlining the significant differences in clinical consequences of the loss of function of this transporter between humans and mice. The markedly different biochemical features of human SLC13A5 and mouse Slc13a5 likely contribute to these differences between humans and mice with regard to the metabolic consequences of the transporter deficiency. The exact molecular mechanisms by which the functional deficiency of the citrate transporter causes epilepsy and impairs neuronal development and function remain to be elucidated, but available literature implicate both dysfunction of GABA (γ-aminobutyrate) signaling and hyperfunction of NMDA (N-methyl-d-aspartate) receptor signaling. Plausible synaptic mechanisms linking loss-of-function mutations in SLC13A5 to epilepsy are discussed.
Keywords: CIC (SLC25A1); GABA; NMDA receptor; NaCT (SLC13A5); cholesterol synthesis; citrate transporter; cytoplasmic citrate; fatty acid synthesis; mitochondrial citrate; neurotransmitters; tricarboxylic acid cycle; zinc.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hodgkinson A. The relation between citric acid and calcium metabolism with particular reference to primary hyper-parathyroidism and idiopathic hypercalcemia. Clin. Sci. 1963;24:167–178. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
