

Polyphosphate granule biogenesis is temporally and functionally tied to cell cycle exit during starvation in Pseudomonas aeruginosa
- PMID: 28265086
- PMCID: PMC5373386
- DOI: 10.1073/pnas.1615575114
Polyphosphate granule biogenesis is temporally and functionally tied to cell cycle exit during starvation in Pseudomonas aeruginosa
Abstract
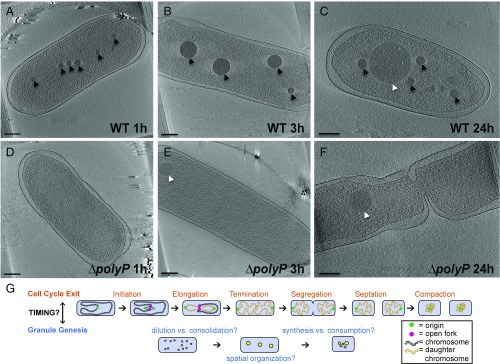
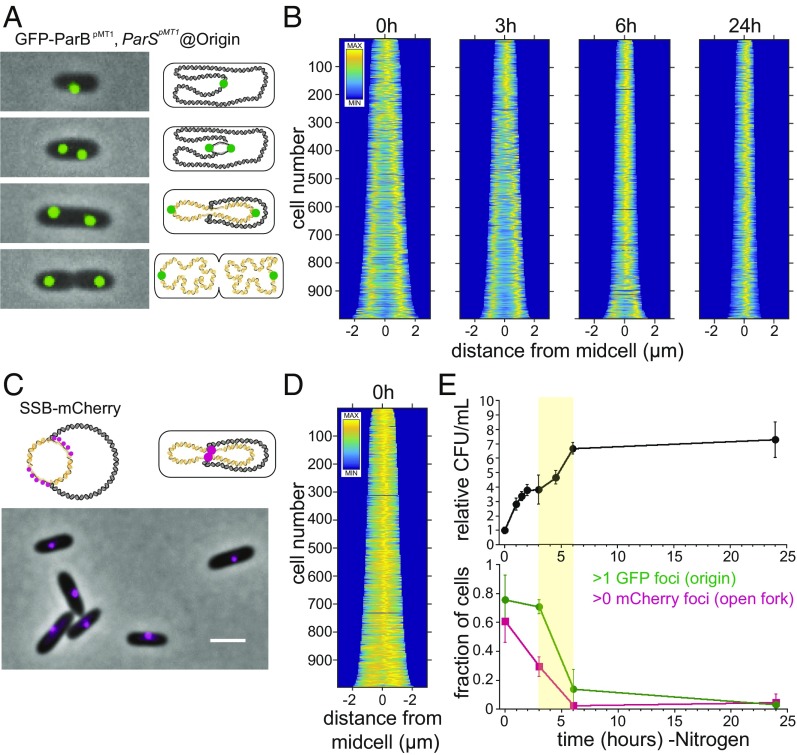
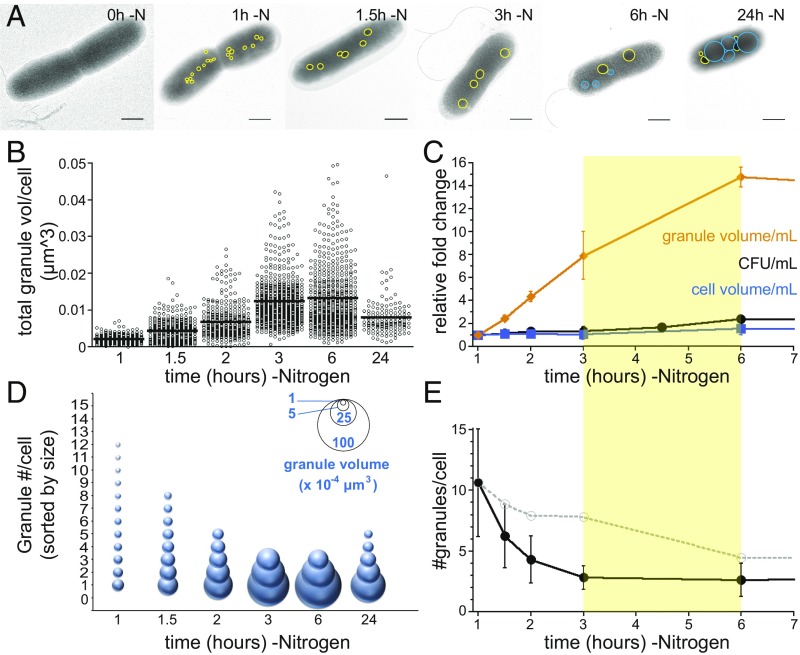
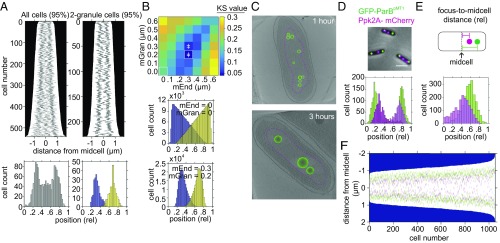
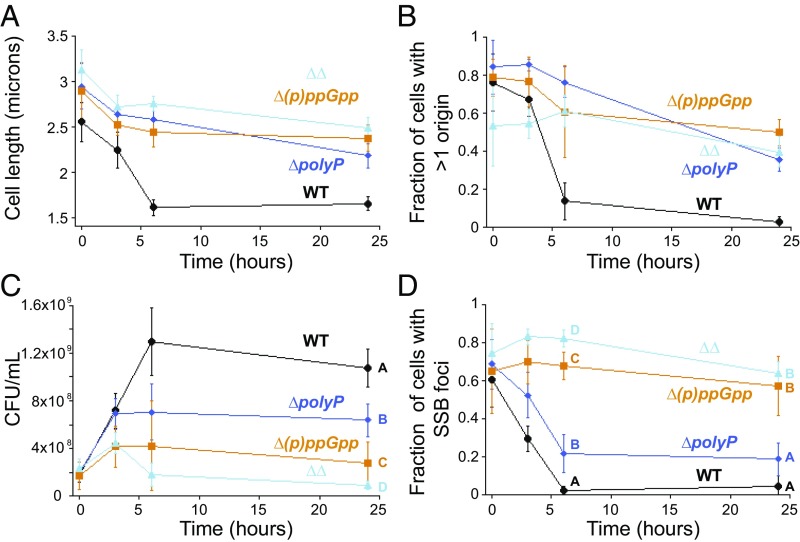
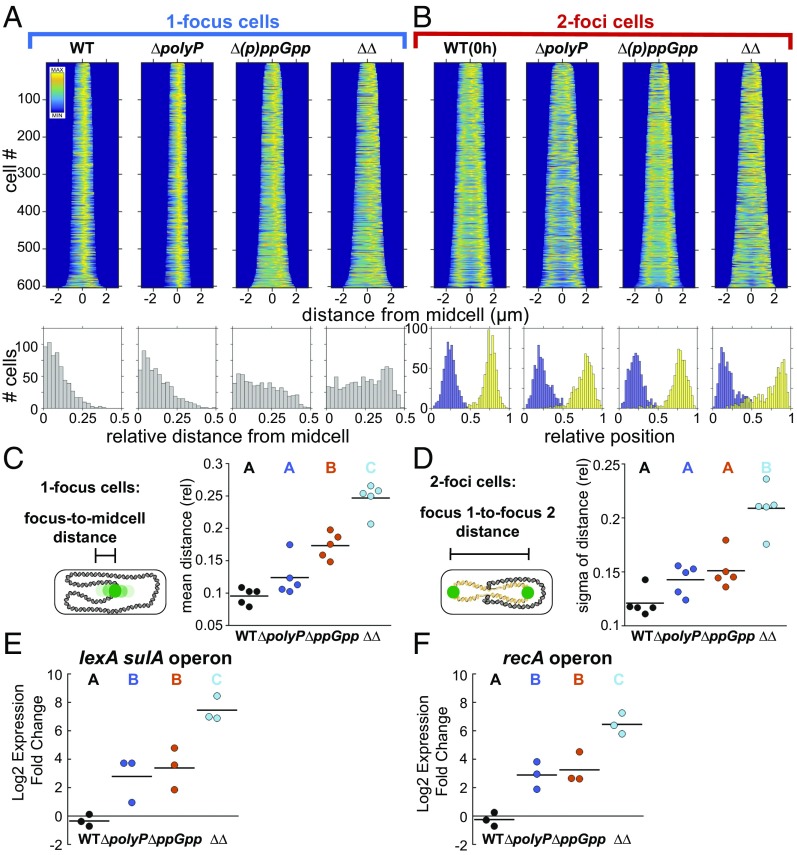
Polyphosphate (polyP) granule biogenesis is an ancient and ubiquitous starvation response in bacteria. Although the ability to make polyP is important for survival during quiescence and resistance to diverse environmental stresses, granule genesis is poorly understood. Using quantitative microscopy at high spatial and temporal resolution, we show that granule genesis in Pseudomonas aeruginosa is tightly organized under nitrogen starvation. Following nucleation as many microgranules throughout the nucleoid, polyP granules consolidate and become transiently spatially organized during cell cycle exit. Between 1 and 3 h after nitrogen starvation, a minority of cells have divided, yet the total granule number per cell decreases, total granule volume per cell dramatically increases, and individual granules grow to occupy diameters as large as ∼200 nm. At their peak, mature granules constitute ∼2% of the total cell volume and are evenly spaced along the long cell axis. Following cell cycle exit, granules initially retain a tight spatial organization, yet their size distribution and spacing relax deeper into starvation. Mutant cells lacking polyP elongate during starvation and contain more than one origin. PolyP promotes cell cycle exit by functioning at a step after DNA replication initiation. Together with the universal starvation alarmone (p)ppGpp, polyP has an additive effect on nucleoid dynamics and organization during starvation. Notably, cell cycle exit is temporally coupled to a net increase in polyP granule biomass, suggesting that net synthesis, rather than consumption of the polymer, is important for the mechanism by which polyP promotes completion of cell cycle exit during starvation.
Keywords: biomineralization; cell cycle; nucleoid; polyphosphate; starvation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Saint-Ruf C, Pesut J, Sopta M, Matic I. Causes and consequences of DNA repair activity modulation during stationary phase in Escherichia coli. Crit Rev Biochem Mol Biol. 2007;42(4):259–270. - PubMed
-
- Minsky A, Shimoni E, Frenkiel-Krispin D. Stress, order and survival. Nat Rev Mol Cell Biol. 2002;3(1):50–60. - PubMed
-
- Frenkiel-Krispin D, Minsky A. Nucleoid organization and the maintenance of DNA integrity in E. coli, B. subtilis and D. radiodurans. J Struct Biol. 2006;156(2):311–319. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
