

Abnormalities in A-to-I RNA editing patterns in CNS injuries correlate with dynamic changes in cell type composition
- PMID: 28266523
- PMCID: PMC5339895
- DOI: 10.1038/srep43421
Abnormalities in A-to-I RNA editing patterns in CNS injuries correlate with dynamic changes in cell type composition
Abstract
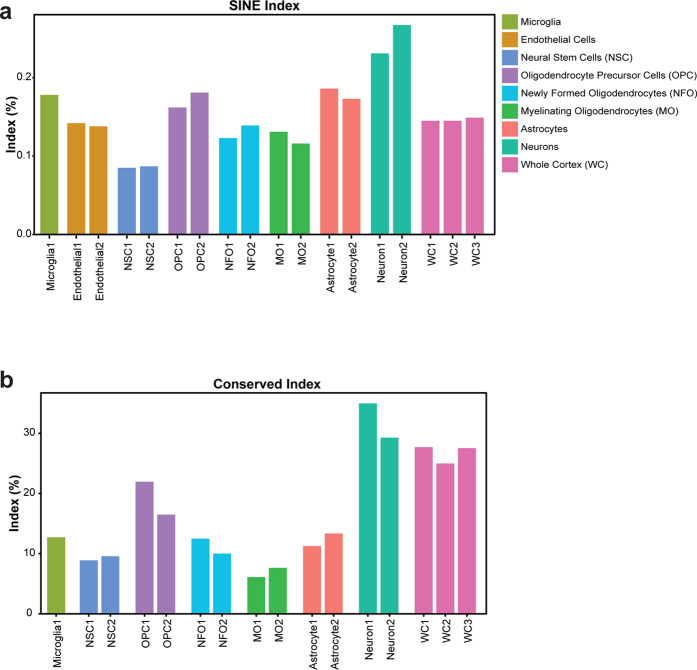
Adenosine to Inosine (A-to-I) RNA editing is a co- or post-transcriptional mechanism that modifies genomically encoded nucleotides at the RNA level. A-to-I RNA editing is abundant in the brain, and altered editing levels have been reported in various neurological pathologies and following spinal cord injury (SCI). The prevailing concept is that the RNA editing process itself is dysregulated by brain pathologies. Here we analyzed recent RNA-seq data, and found that, except for few mammalian conserved editing sites, editing is significantly higher in neurons than in other cell populations of the brain. We studied A-to-I RNA editing in stab wound injury (SWI) and SCI models and showed that the apparent under-editing observed after injury correlates with an approximately 20% reduction in the relative density of neurons, due to cell death and immune cell infiltration that may account for the observed under-editing. Studies of neuronal and astrocyte cultures and a computational analysis of SCI RNA-seq data further supported the possibility that a reduction in neuronal density is responsible for alterations in the tissue-wide editing patterns upon injury. Thus, our data suggest that the case for a mechanistic linkage between A-to-I RNA editing and brain pathologies should be revisited.
Conflict of interest statement
The authors declare no competing financial interests.
Figures





Similar articles
-
Reactive astrocytes undergo M1 microglia/macrohpages-induced necroptosis in spinal cord injury.Mol Neurodegener. 2016 Feb 3;11:14. doi: 10.1186/s13024-016-0081-8. Mol Neurodegener. 2016. PMID: 26842216 Free PMC article.
-
Spatiotemporal expression of FOXA1 correlates with reactive gliosis after spinal cord injury.Neuropeptides. 2017 Dec;66:36-44. doi: 10.1016/j.npep.2017.08.002. Epub 2017 Aug 18. Neuropeptides. 2017. PMID: 28844448
-
Ablation of the transcription factors E2F1-2 limits neuroinflammation and associated neurological deficits after contusive spinal cord injury.Cell Cycle. 2015;14(23):3698-712. doi: 10.1080/15384101.2015.1104436. Cell Cycle. 2015. PMID: 26505089 Free PMC article.
-
Recent Advances in Adenosine-to-Inosine RNA Editing in Cancer.Cancer Treat Res. 2023;190:143-179. doi: 10.1007/978-3-031-45654-1_5. Cancer Treat Res. 2023. PMID: 38113001 Review.
-
Glial-Neuronal Interactions in Pathogenesis and Treatment of Spinal Cord Injury.Int J Mol Sci. 2021 Dec 17;22(24):13577. doi: 10.3390/ijms222413577. Int J Mol Sci. 2021. PMID: 34948371 Free PMC article. Review.
Cited by
-
A-to-I RNA editing in the rat brain is age-dependent, region-specific and sensitive to environmental stress across generations.BMC Genomics. 2018 Jan 8;19(1):28. doi: 10.1186/s12864-017-4409-8. BMC Genomics. 2018. PMID: 29310578 Free PMC article.
-
Decreased A-to-I RNA editing as a source of keratinocytes' dsRNA in psoriasis.RNA. 2018 Jun;24(6):828-840. doi: 10.1261/rna.064659.117. Epub 2018 Mar 28. RNA. 2018. PMID: 29592874 Free PMC article.
-
Recommendations for detection, validation, and evaluation of RNA editing events in cardiovascular and neurological/neurodegenerative diseases.Mol Ther Nucleic Acids. 2023 Dec 5;35(1):102085. doi: 10.1016/j.omtn.2023.102085. eCollection 2024 Mar 12. Mol Ther Nucleic Acids. 2023. PMID: 38192612 Free PMC article. Review.
-
Distant coupling between RNA editing and alternative splicing of the osmosensitive cation channel Tmem63b.J Biol Chem. 2020 Dec 25;295(52):18199-18212. doi: 10.1074/jbc.RA120.016049. Epub 2020 Oct 25. J Biol Chem. 2020. PMID: 33100268 Free PMC article.
-
Epitranscriptomic Modifications Modulate Normal and Pathological Functions in CNS.Transl Stroke Res. 2022 Feb;13(1):1-11. doi: 10.1007/s12975-021-00927-z. Epub 2021 Jul 5. Transl Stroke Res. 2022. PMID: 34224107 Free PMC article. Review.
References
-
- Hough R. F. & Bass B. L. Purification of the Xenopus laevis double-stranded RNA adenosine deaminase. J. Biol. Chem. 269, 9933–9 (1994). - PubMed
-
- Melcher T. et al.. A mammalian RNA editing enzyme. Nature 379, 460–4 (1996). - PubMed
-
- Melcher T. et al.. RED2, a brain-specific member of the RNA-specific adenosine deaminase family. J. Biol. Chem. 271, 31795–8 (1996). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
