

Commensal gut bacteria modulate phosphorylation-dependent PPARγ transcriptional activity in human intestinal epithelial cells
- PMID: 28266623
- PMCID: PMC5339702
- DOI: 10.1038/srep43199
Commensal gut bacteria modulate phosphorylation-dependent PPARγ transcriptional activity in human intestinal epithelial cells
Abstract
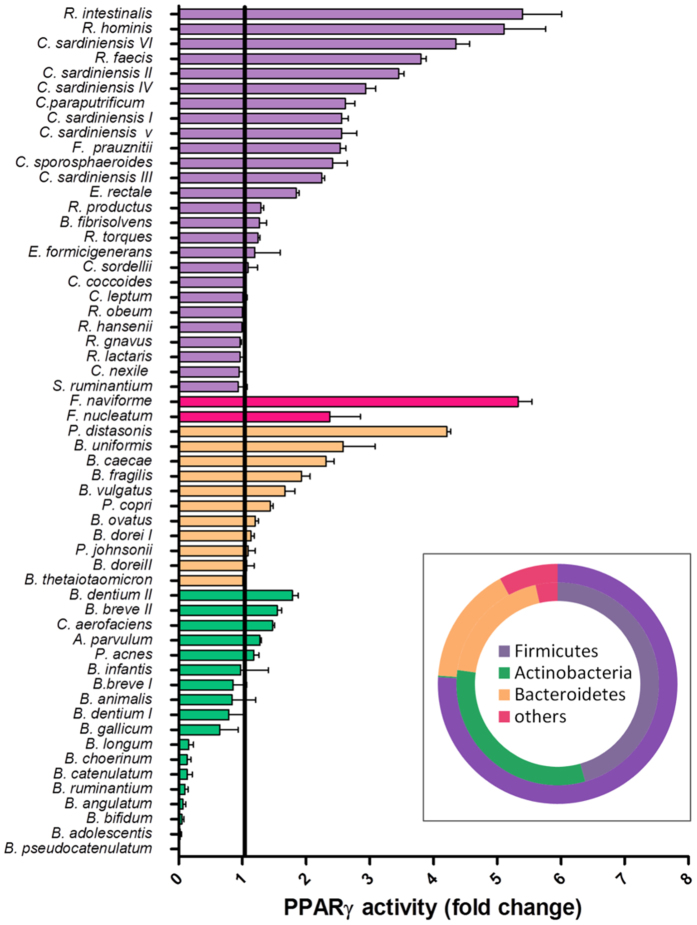
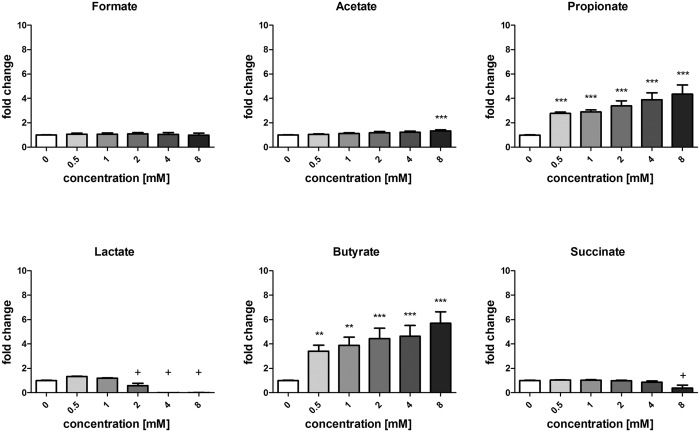
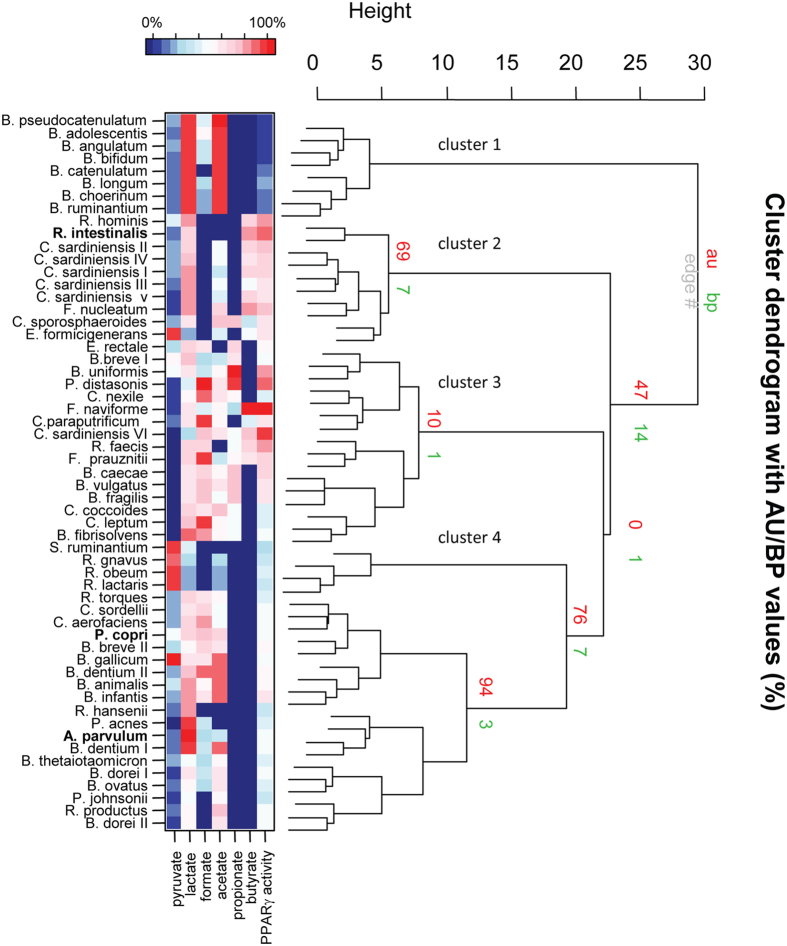
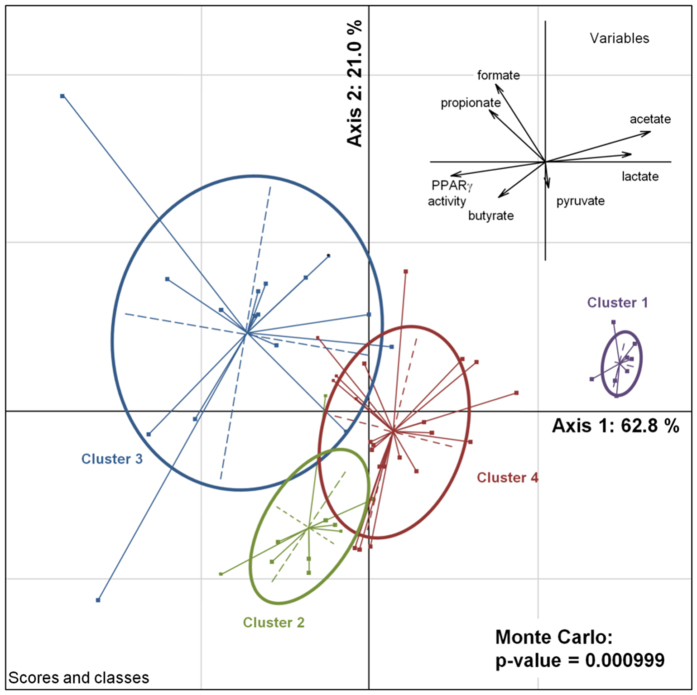
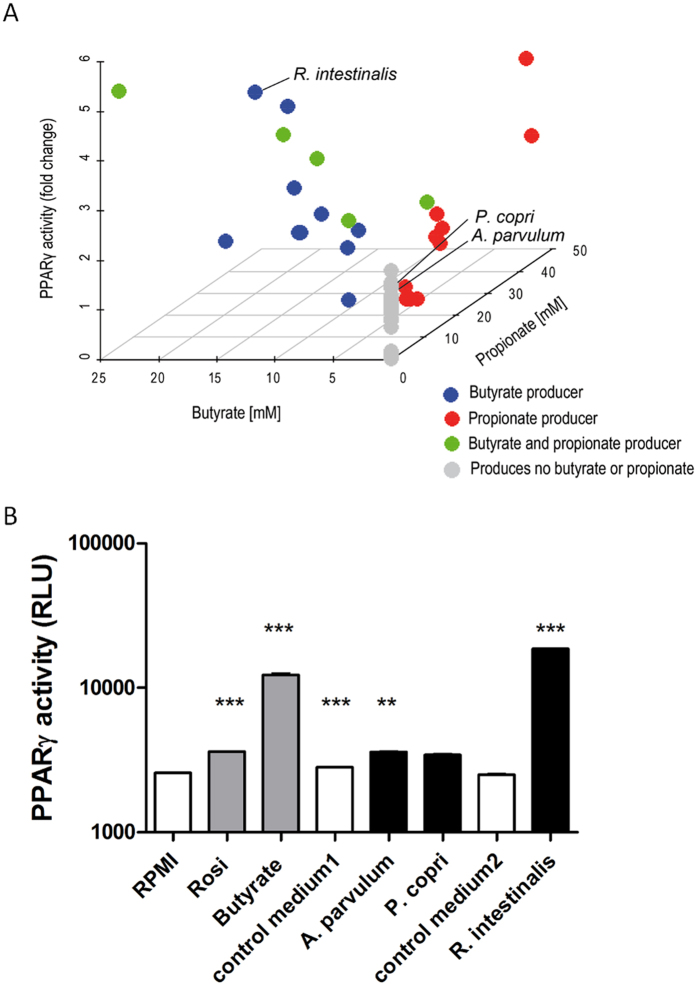
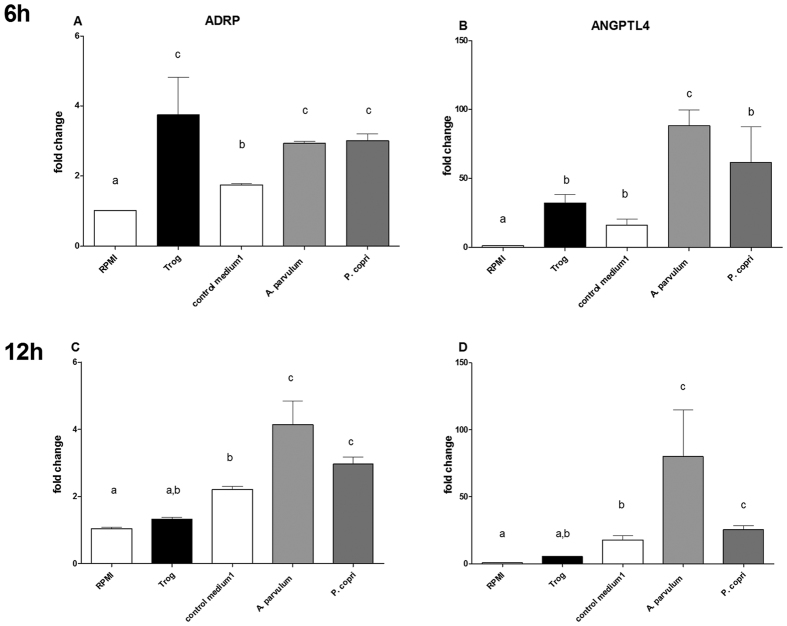
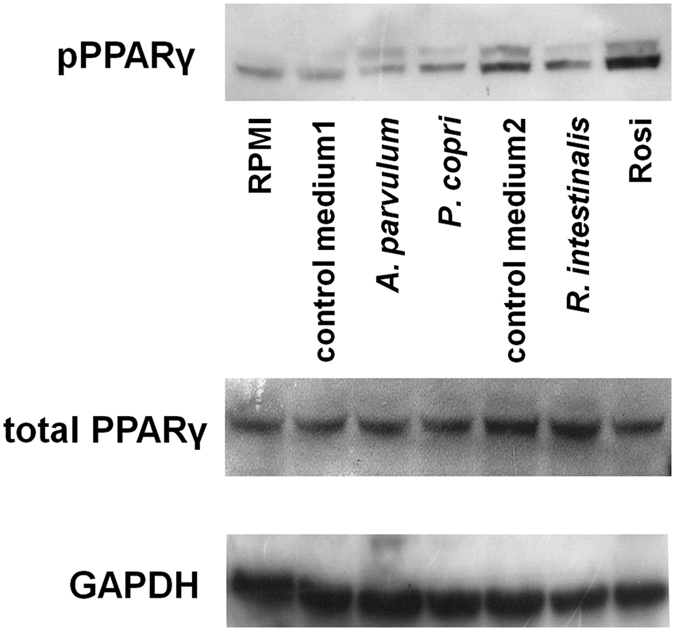
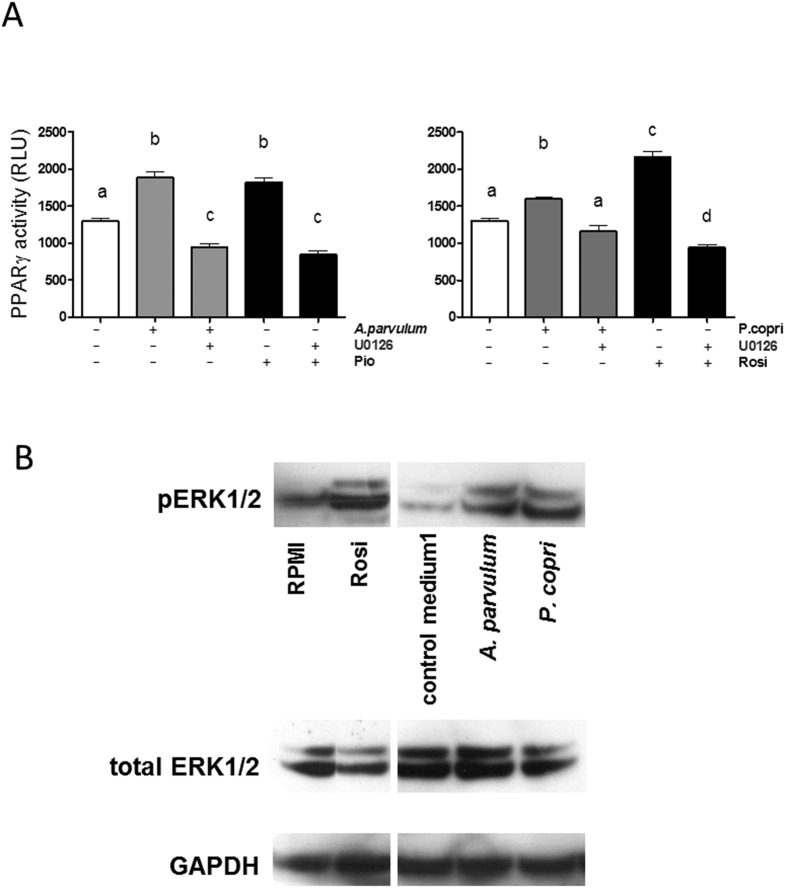
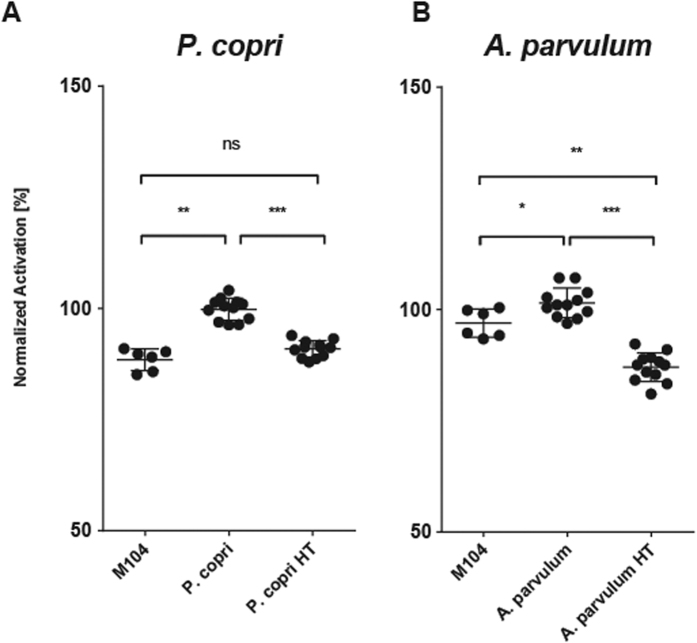
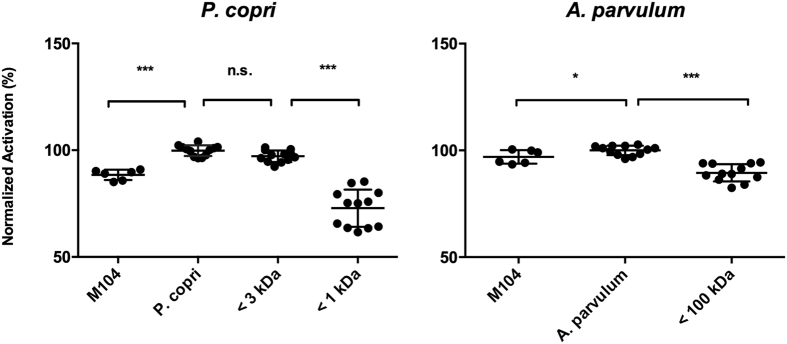
In healthy subjects, the intestinal microbiota interacts with the host's epithelium, regulating gene expression to the benefit of both, host and microbiota. The underlying mechanisms remain poorly understood, however. Although many gut bacteria are not yet cultured, constantly growing culture collections have been established. We selected 57 representative commensal bacterial strains to study bacteria-host interactions, focusing on PPARγ, a key nuclear receptor in colonocytes linking metabolism and inflammation to the microbiota. Conditioned media (CM) were harvested from anaerobic cultures and assessed for their ability to modulate PPARγ using a reporter cell line. Activation of PPARγ transcriptional activity was linked to the presence of butyrate and propionate, two of the main metabolites of intestinal bacteria. Interestingly, some stimulatory CMs were devoid of these metabolites. A Prevotella and an Atopobium strain were chosen for further study, and shown to up-regulate two PPARγ-target genes, ANGPTL4 and ADRP. The molecular mechanisms of these activations involved the phosphorylation of PPARγ through ERK1/2. The responsible metabolites were shown to be heat sensitive but markedly diverged in size, emphasizing the diversity of bioactive compounds found in the intestine. Here we describe different mechanisms by which single intestinal bacteria can directly impact their host's health through transcriptional regulation.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Sekirov I., Russell S. L., Antunes L. C. M. & Finlay B. B. Gut Microbiota in Health and Disease. Physiol. Rev. 90, 859–904 (2010). - PubMed
-
- Cerf-Bensussan N. & Gaboriau-Routhiau V. The immune system and the gut microbiota: friends or foes? Nat. Rev. Immunol. 10, 735–44 (2010). - PubMed
-
- Cummings J. H. Microbial Digestion of Complex Carbohydrates in Man. Proc. Nutr. Soc. 43, 35–44 (1984). - PubMed
-
- Albert M. J., Mathan V. I. & Baker S. J. Vitamin B12 synthesis by human small intestinal bacteria. Nature 283, 781–782 (1980). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
