

Single-molecule studies reveal reciprocating of WRN helicase core along ssDNA during DNA unwinding
- PMID: 28266653
- PMCID: PMC5339710
- DOI: 10.1038/srep43954
Single-molecule studies reveal reciprocating of WRN helicase core along ssDNA during DNA unwinding
Abstract
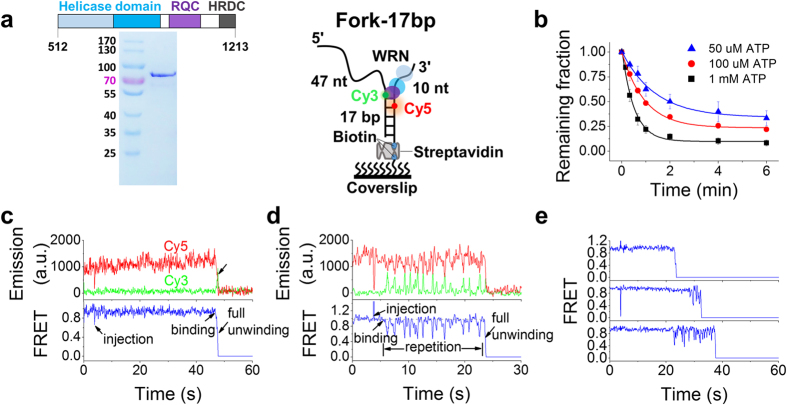
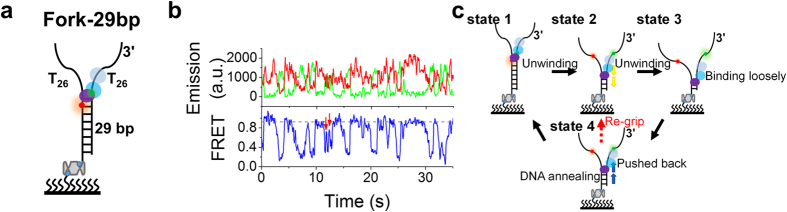
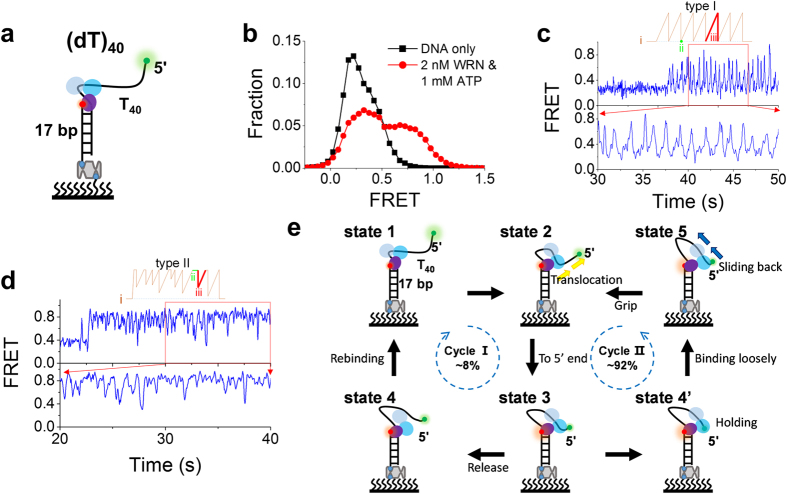
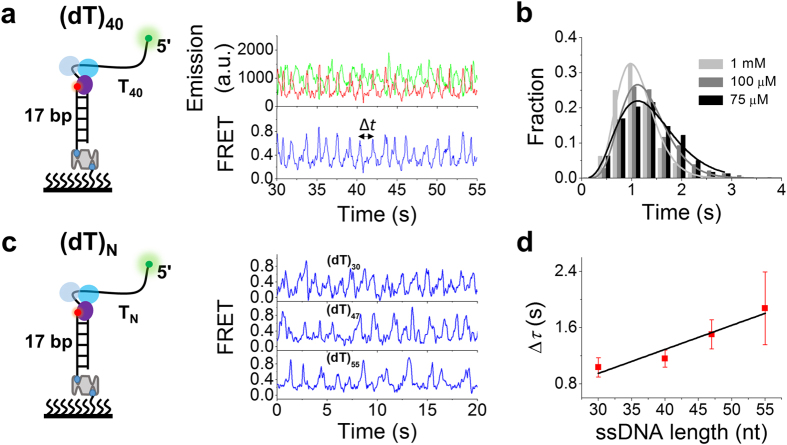
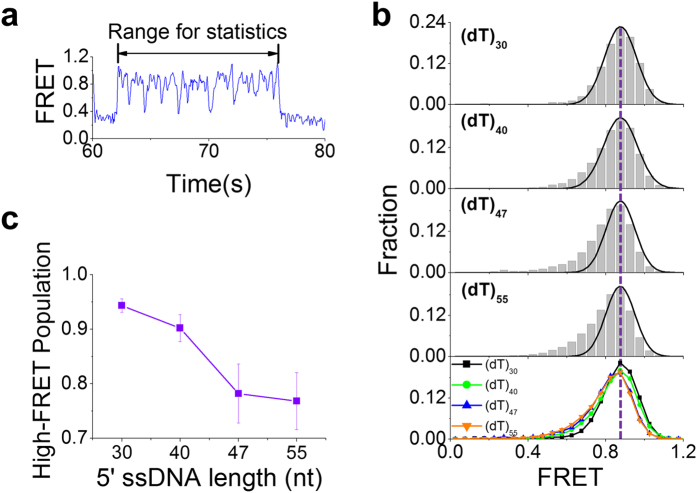
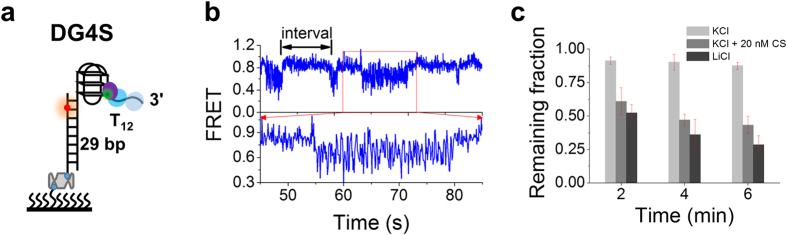
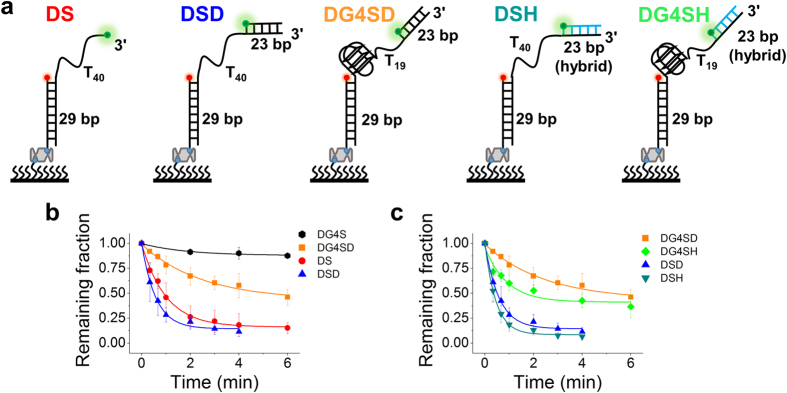
Werner syndrome is caused by mutations in the WRN gene encoding WRN helicase. A knowledge of WRN helicase's DNA unwinding mechanism in vitro is helpful for predicting its behaviors in vivo, and then understanding their biological functions. In the present study, for deeply understanding the DNA unwinding mechanism of WRN, we comprehensively characterized the DNA unwinding properties of chicken WRN helicase core in details, by taking advantages of single-molecule fluorescence resonance energy transfer (smFRET) method. We showed that WRN exhibits repetitive DNA unwinding and translocation behaviors on different DNA structures, including forked, overhanging and G-quadruplex-containing DNAs with an apparently limited unwinding processivity. It was further revealed that the repetitive behaviors were caused by reciprocating of WRN along the same ssDNA, rather than by complete dissociation from and rebinding to substrates or by strand switching. The present study sheds new light on the mechanism for WRN functioning.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Singleton M. R., Dillingham M. S. & Wigley D. B. Structure and mechanism of helicases and nucleic acid translocases. Annu. Rev. Biochem. 76, 23–50 (2007). - PubMed
-
- van Brabant A. J., Stan R. & Ellis N. A. DNA helicases, genomic instability, and human genetic disease. Annu. Rev. Genomics Hum. Genet. 1, 409–459 (2000). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
