

The DRF motif of CXCR6 as chemokine receptor adaptation to adhesion
- PMID: 28267793
- PMCID: PMC5340378
- DOI: 10.1371/journal.pone.0173486
The DRF motif of CXCR6 as chemokine receptor adaptation to adhesion
Abstract
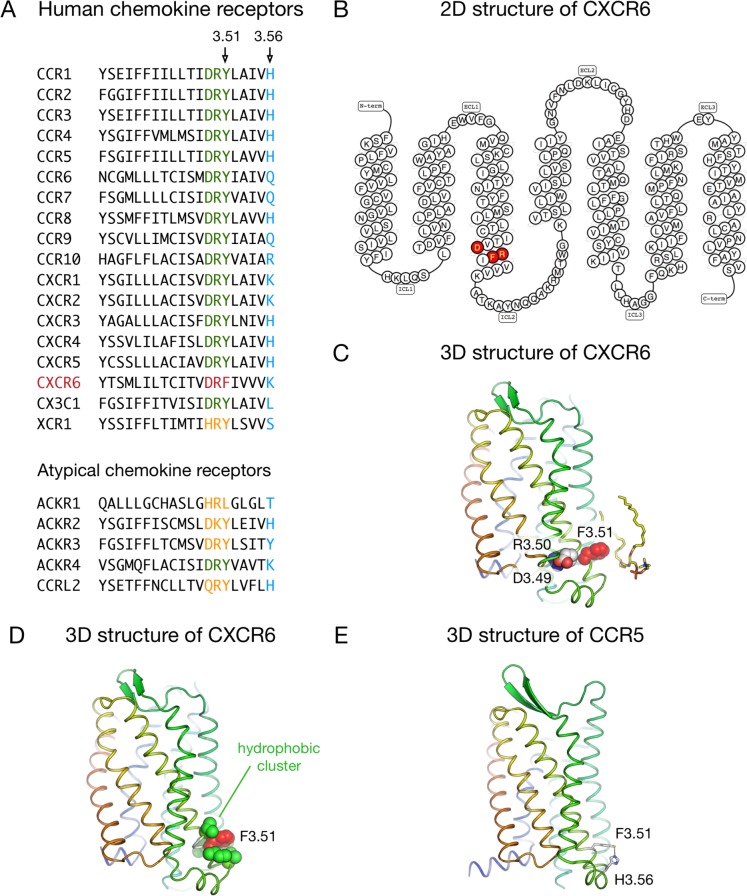
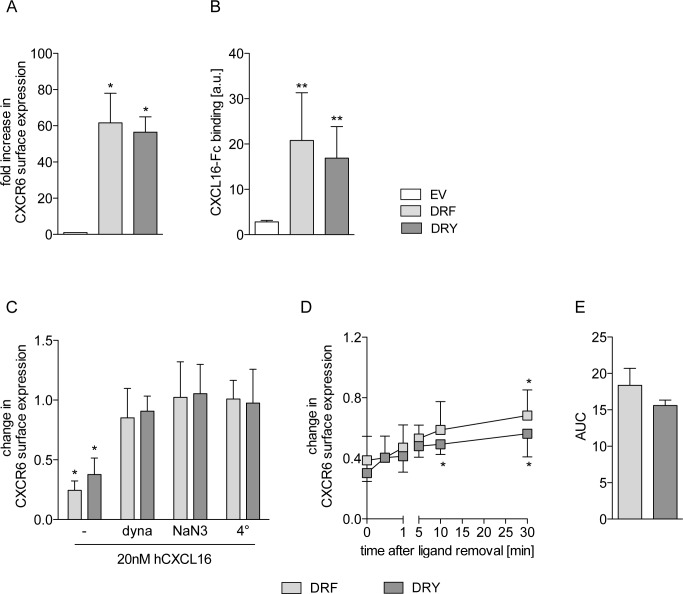
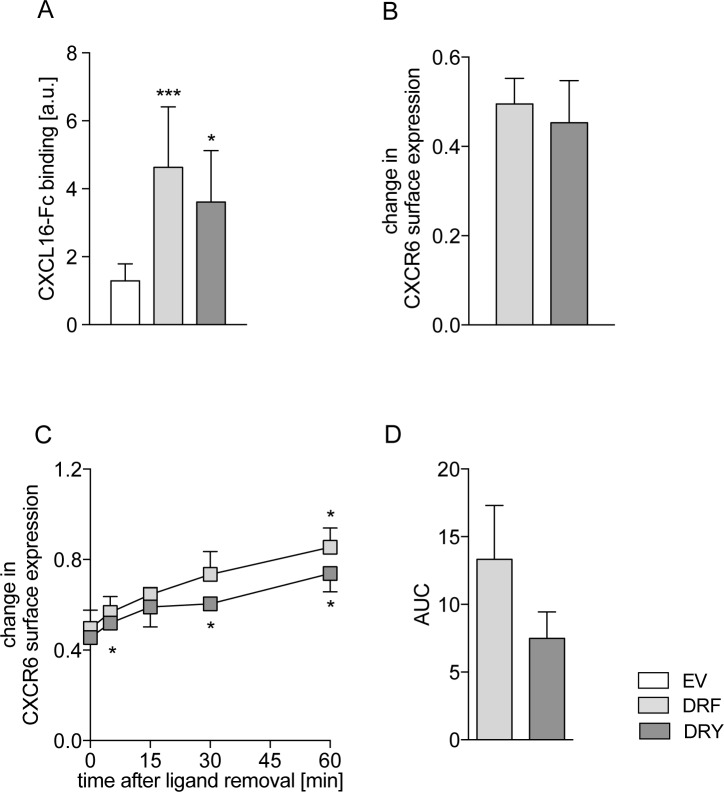
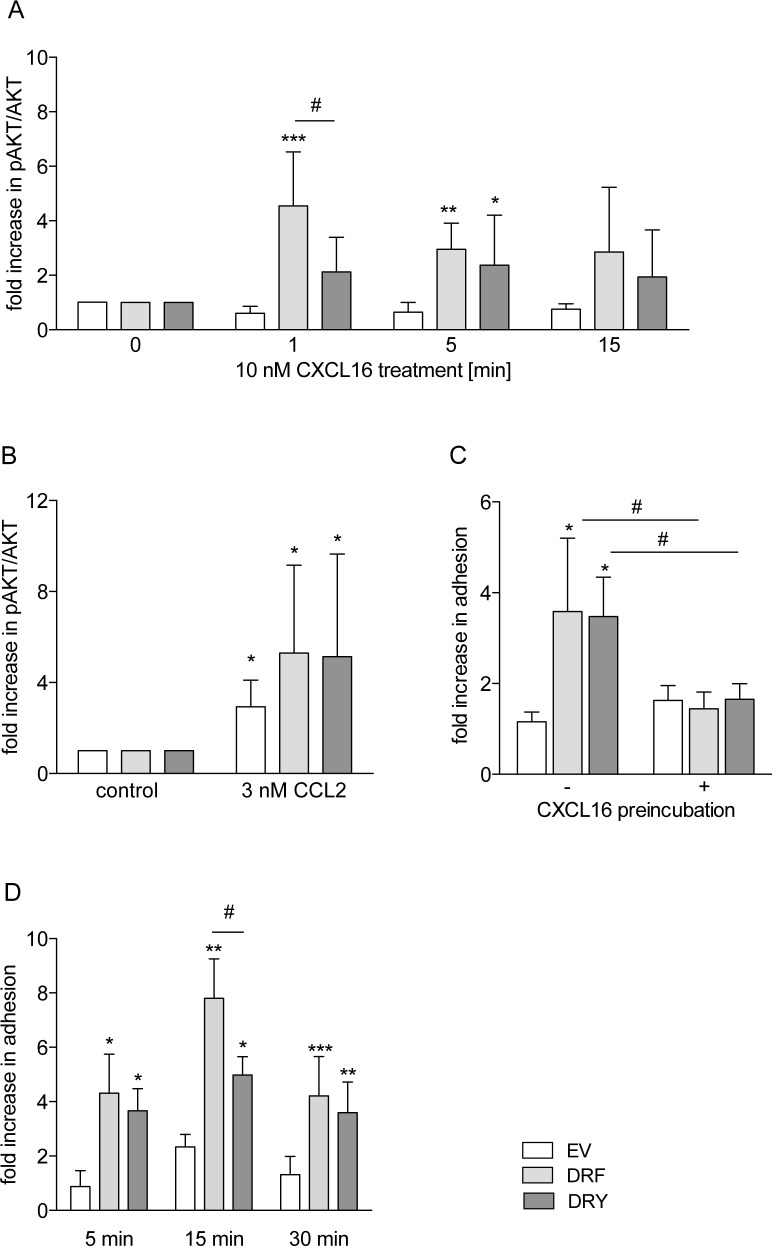
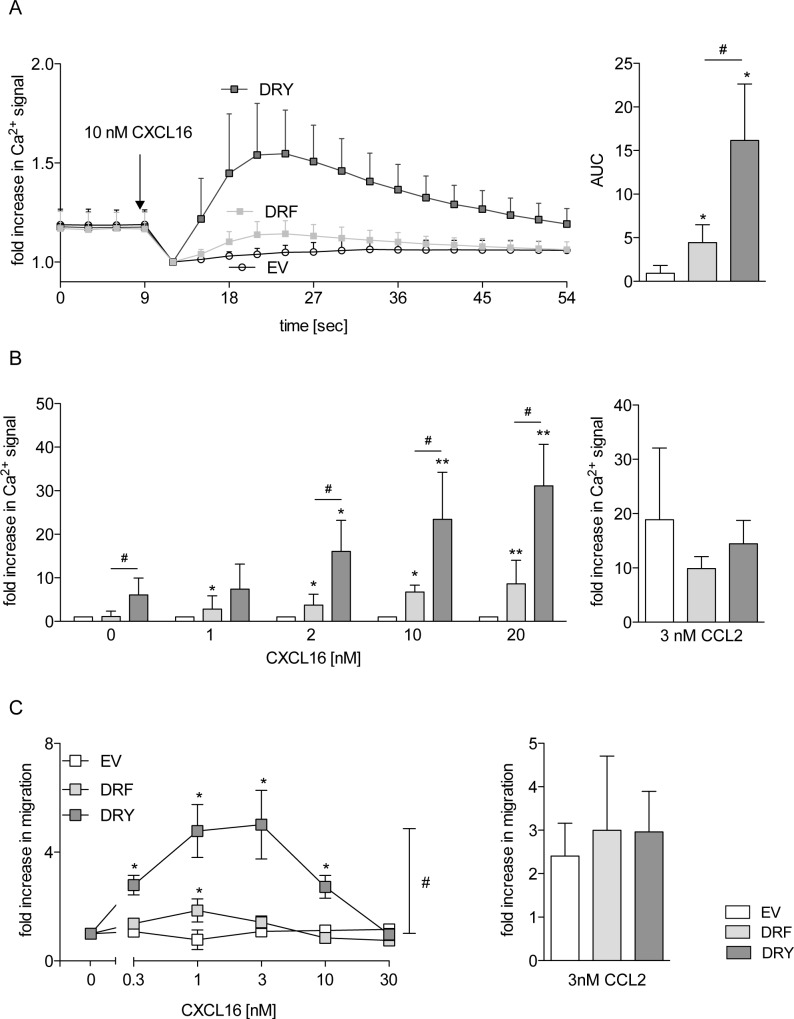
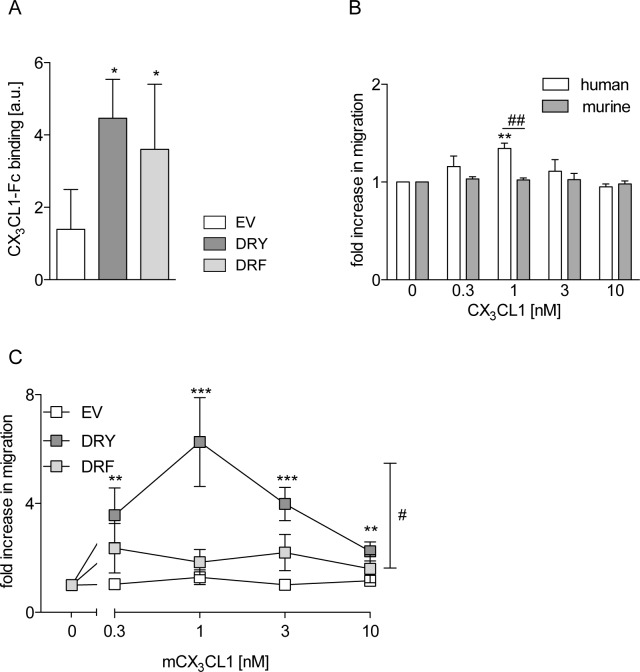
The CXC-chemokine receptor 6 (CXCR6) is a class A GTP-binding protein-coupled receptor (GPCRs) that mediates adhesion of leukocytes by interacting with the transmembrane cell surface-expressed chemokine ligand 16 (CXCL16), and also regulates leukocyte migration by interacting with the soluble shed variant of CXCL16. In contrast to virtually all other chemokine receptors with chemotactic activity, CXCR6 carries a DRF motif instead of the typical DRY motif as a key element in receptor activation and G protein coupling. In this work, modeling analyses revealed that the phenylalanine F3.51 in CXCR6 might have impact on intramolecular interactions including hydrogen bonds by this possibly changing receptor function. Initial investigations with embryonic kidney HEK293 cells and further studies with monocytic THP-1 cells showed that mutation of DRF into DRY does not influence ligand binding, receptor internalization, receptor recycling, and protein kinase B (AKT) signaling. Adhesion was slightly decreased in a time-dependent manner. However, CXCL16-induced calcium signaling and migration were increased. Vice versa, when the DRY motif of the related receptor CX3CR1 was mutated into DRF the migratory response towards CX3CL1 was diminished, indicating that the presence of a DRF motif generally impairs chemotaxis in chemokine receptors. Transmembrane and soluble CXCL16 play divergent roles in homeostasis, inflammation, and cancer, which can be beneficial or detrimental. Therefore, the DRF motif of CXCR6 may display a receptor adaptation allowing adhesion and cell retention by transmembrane CXCL16 but reducing the chemotactic response to soluble CXCL16. This adaptation may avoid permanent or uncontrolled recruitment of inflammatory cells as well as cancer metastasis.
Conflict of interest statement
Figures
Similar articles
-
Selectivity in the Use of Gi/o Proteins Is Determined by the DRF Motif in CXCR6 and Is Cell-Type Specific.Mol Pharmacol. 2015 Nov;88(5):894-910. doi: 10.1124/mol.115.099960. Epub 2015 Aug 27. Mol Pharmacol. 2015. PMID: 26316539 Free PMC article.
-
Homocysteine up-regulates vascular transmembrane chemokine CXCL16 and induces CXCR6+ lymphocyte recruitment in vitro and in vivo.J Cell Mol Med. 2008 Sep-Oct;12(5A):1700-9. doi: 10.1111/j.1582-4934.2008.00223.x. Epub 2008 Jan 11. J Cell Mol Med. 2008. PMID: 18194461 Free PMC article.
-
The chemokine receptor CXCR6 and its ligand CXCL16 are expressed in carcinomas and inhibit proliferation.Cancer Res. 2008 Jun 15;68(12):4701-8. doi: 10.1158/0008-5472.CAN-08-0482. Cancer Res. 2008. Retraction in: Cancer Res. 2011 Feb 1;71(3):1196. doi: 10.1158/0008-5472.CAN-10-4378. PMID: 18559516 Retracted.
-
CXCR6/CXCL16 functions as a regulator in metastasis and progression of cancer.Biochim Biophys Acta. 2010 Aug;1806(1):42-9. doi: 10.1016/j.bbcan.2010.01.004. Epub 2010 Feb 1. Biochim Biophys Acta. 2010. PMID: 20122997 Review.
-
The role of the CXCR6/CXCL16 axis in the pathogenesis of fibrotic disease.Int Immunopharmacol. 2024 May 10;132:112015. doi: 10.1016/j.intimp.2024.112015. Epub 2024 Apr 11. Int Immunopharmacol. 2024. PMID: 38608478 Review.
Cited by
-
CXCL16/CXCR6 Axis in Adipocytes Differentiated from Human Adipose Derived Mesenchymal Stem Cells Regulates Macrophage Polarization.Cells. 2021 Dec 3;10(12):3410. doi: 10.3390/cells10123410. Cells. 2021. PMID: 34943917 Free PMC article.
-
Cryo-EM structure of G-protein-coupled receptor GPR17 in complex with inhibitory G protein.MedComm (2020). 2022 Sep 10;3(4):e159. doi: 10.1002/mco2.159. eCollection 2022 Dec. MedComm (2020). 2022. PMID: 36105372 Free PMC article.
-
Behavior of Chemokine Receptor 6 (CXCR6) in Complex with CXCL16 Soluble form Chemokine by Molecular Dynamic Simulations: General Protein‒Ligand Interaction Model and 3D-QSAR Studies of Synthetic Antagonists.Life (Basel). 2021 Apr 15;11(4):346. doi: 10.3390/life11040346. Life (Basel). 2021. PMID: 33920834 Free PMC article.
-
CXCR6 positions cytotoxic T cells to receive critical survival signals in the tumor microenvironment.Cell. 2021 Aug 19;184(17):4512-4530.e22. doi: 10.1016/j.cell.2021.07.015. Epub 2021 Aug 2. Cell. 2021. PMID: 34343496 Free PMC article.
-
CXCL1: Gene, Promoter, Regulation of Expression, mRNA Stability, Regulation of Activity in the Intercellular Space.Int J Mol Sci. 2022 Jan 12;23(2):792. doi: 10.3390/ijms23020792. Int J Mol Sci. 2022. PMID: 35054978 Free PMC article. Review.
References
-
- Latta M, Mohan K, Issekutz TB. CXCR6 is expressed on T cells in both T helper type 1 (Th1) inflammation and allergen-induced Th2 lung inflammation but is only a weak mediator of chemotaxis. Immunology. 2007. August;121(4):555–64. Epub 2007/04/18. eng. 10.1111/j.1365-2567.2007.02603.x - DOI - PMC - PubMed
-
- Chandrasekar B, Bysani S, Mummidi S. CXCL16 signals via Gi, phosphatidylinositol 3-kinase, Akt, I kappa B kinase, and nuclear factor-kappa B and induces cell-cell adhesion and aortic smooth muscle cell proliferation. J Biol Chem. 2004. January 30;279(5):3188–96. Epub 2003/11/20. eng. 10.1074/jbc.M311660200 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
