

Cbx3 maintains lineage specificity during neural differentiation
- PMID: 28270516
- PMCID: PMC5358721
- DOI: 10.1101/gad.292169.116
Cbx3 maintains lineage specificity during neural differentiation
Abstract
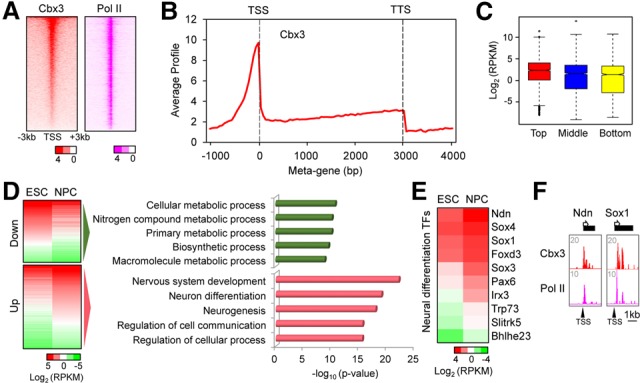
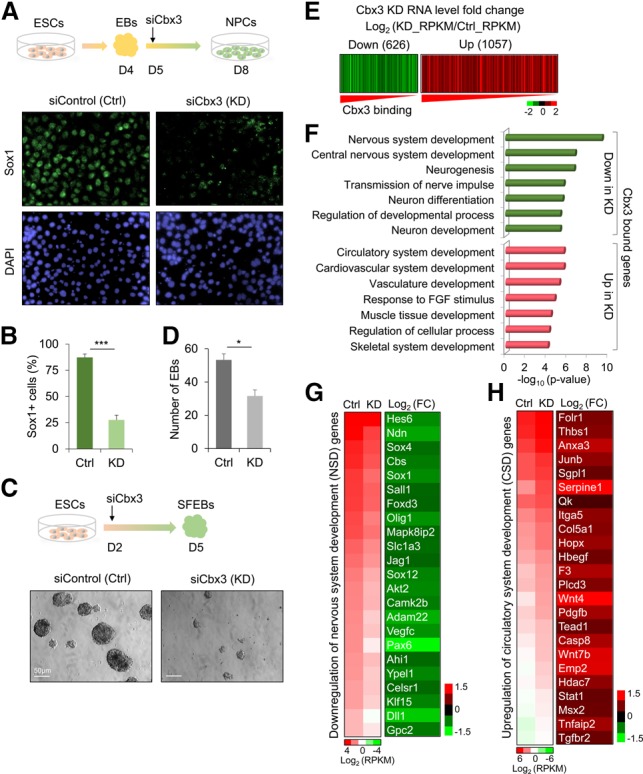
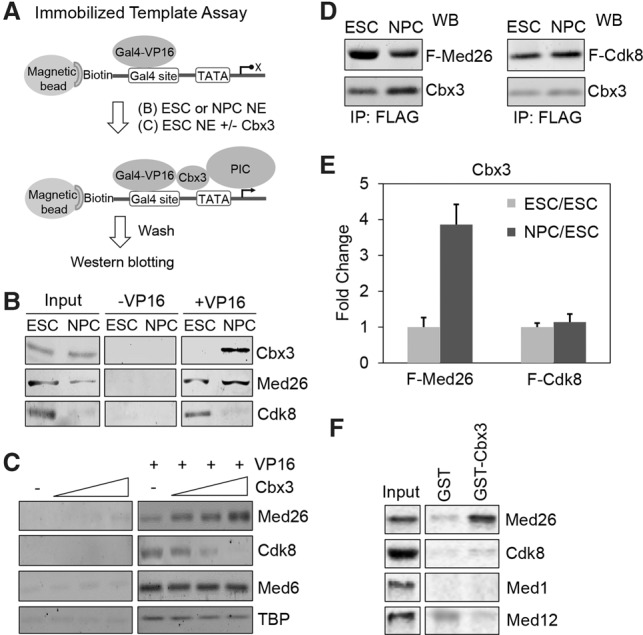
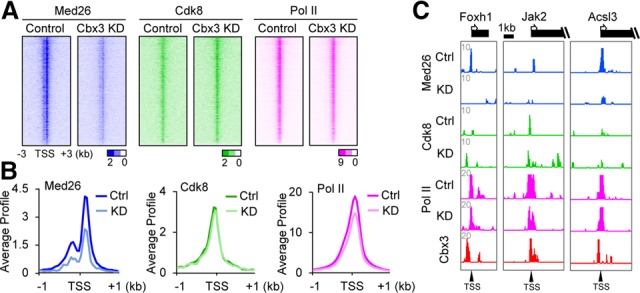
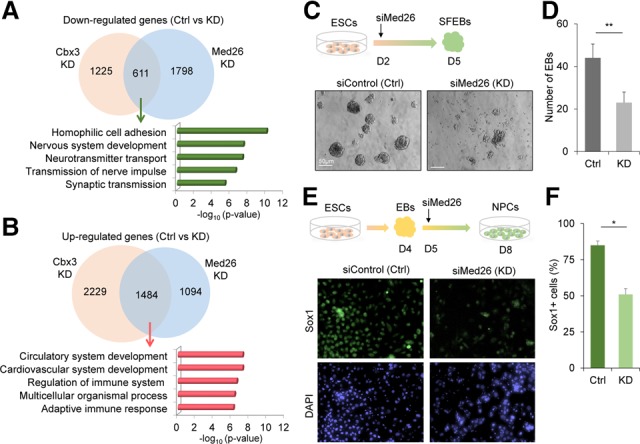
Chromobox homolog 3 (Cbx3/heterochromatin protein 1γ [HP1γ]) stimulates cell differentiation, but its mechanism is unknown. We found that Cbx3 binds to gene promoters upon differentiation of murine embryonic stem cells (ESCs) to neural progenitor cells (NPCs) and recruits the Mediator subunit Med26. RNAi knockdown of either Cbx3 or Med26 inhibits neural differentiation while up-regulating genes involved in mesodermal lineage decisions. Thus, Cbx3 and Med26 together ensure the fidelity of lineage specification by enhancing the expression of neural genes and down-regulating genes specific to alternative fates.
Keywords: Cbx3; Med26; embryonic stem cell; mesoderm; neural precursor; preinitiation complex.
© 2017 Huang et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Beard C, Hochedlinger K, Plath K, Wutz A, Jaenisch R. 2006. Efficient method to generate single-copy transgenic mice by site-specific integration in embryonic stem cells. Genesis 44: 23–28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous