

Mild Cognitive Impairment Is Characterized by Deficient Brainstem and Cortical Representations of Speech
- PMID: 28270574
- PMCID: PMC6596919
- DOI: 10.1523/JNEUROSCI.3700-16.2017
Mild Cognitive Impairment Is Characterized by Deficient Brainstem and Cortical Representations of Speech
Abstract
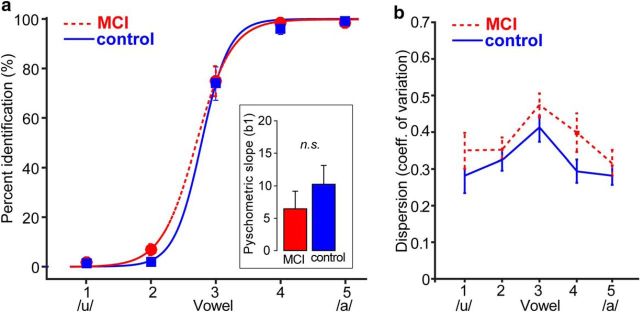
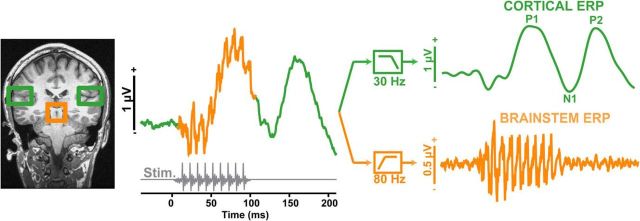
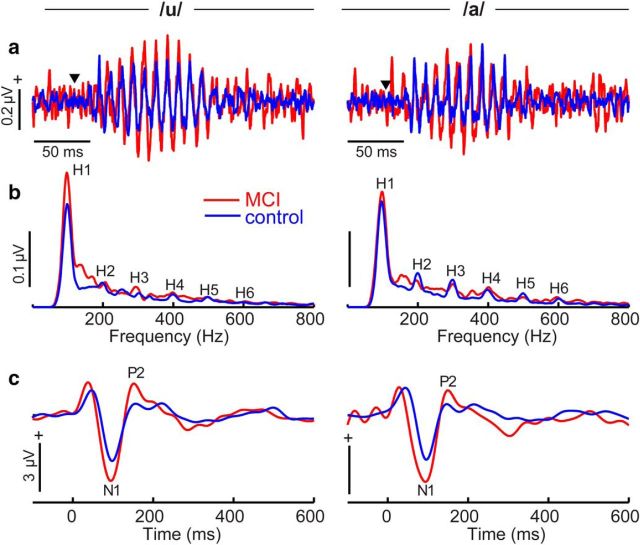
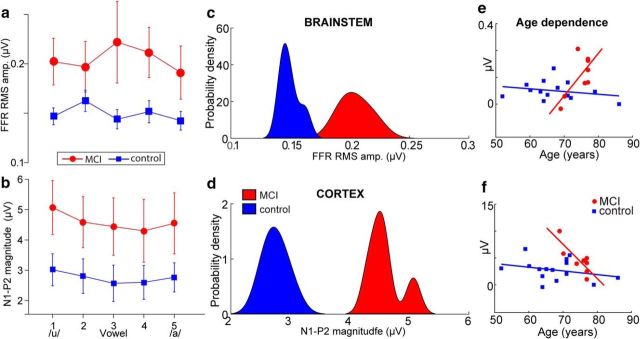
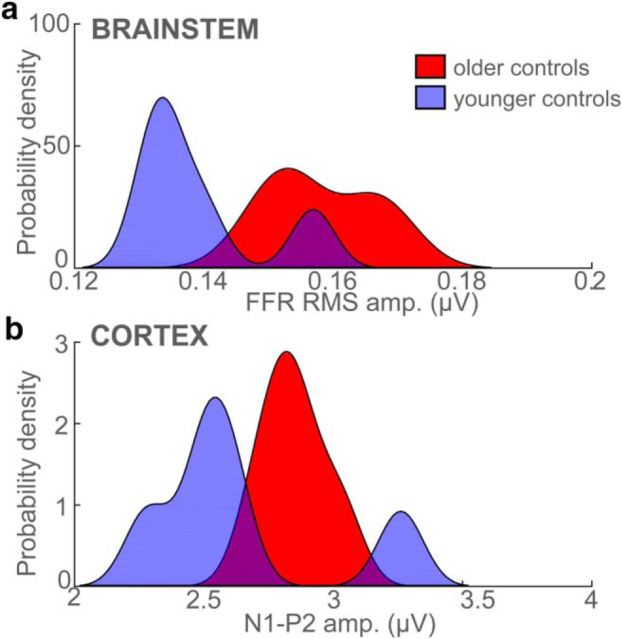
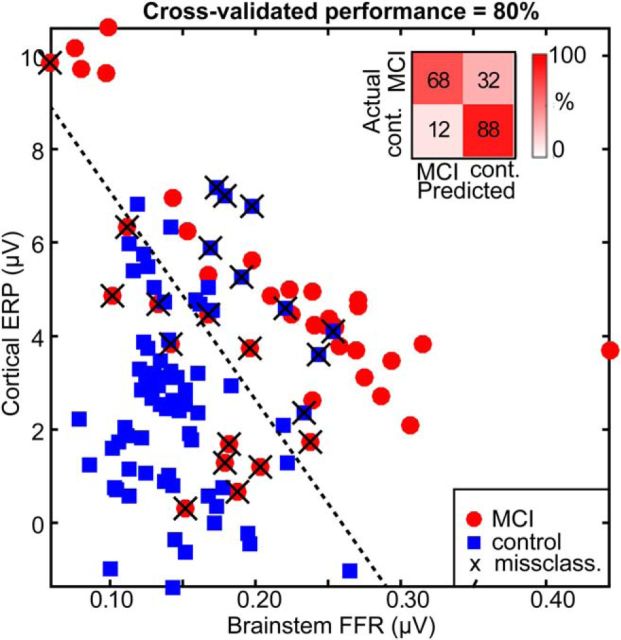
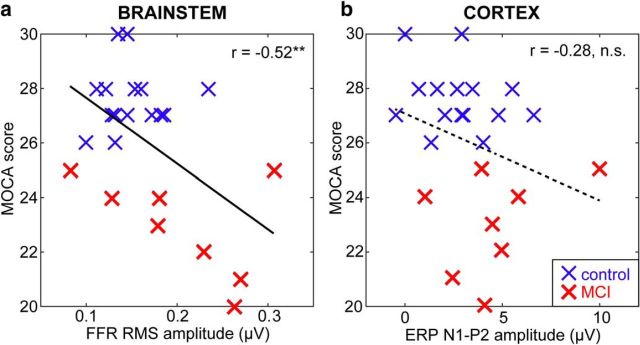
Mild cognitive impairment (MCI) is recognized as a transitional phase in the progression toward more severe forms of dementia and is an early precursor to Alzheimer's disease. Previous neuroimaging studies reveal that MCI is associated with aberrant sensory-perceptual processing in cortical brain regions subserving auditory and language function. However, whether the pathophysiology of MCI extends to speech processing before conscious awareness (brainstem) is unknown. Using a novel electrophysiological approach, we recorded both brainstem and cortical speech-evoked brain event-related potentials (ERPs) in older, hearing-matched human listeners who did and did not present with subtle cognitive impairment revealed through behavioral neuropsychological testing. We found that MCI was associated with changes in neural speech processing characterized as hypersensitivity (larger) brainstem and cortical speech encoding in MCI compared with controls in the absence of any perceptual speech deficits. Group differences also interacted with age differentially across the auditory pathway; brainstem responses became larger and cortical ERPs smaller with advancing age. Multivariate classification revealed that dual brainstem-cortical speech activity correctly identified MCI listeners with 80% accuracy, suggesting its application as a biomarker of early cognitive decline. Brainstem responses were also a more robust predictor of individuals' MCI severity than cortical activity. Our findings suggest that MCI is associated with poorer encoding and transfer of speech signals between functional levels of the auditory system and advance the pathophysiological understanding of cognitive aging by identifying subcortical deficits in auditory sensory processing mere milliseconds (<10 ms) after sound onset and before the emergence of perceptual speech deficits.SIGNIFICANCE STATEMENT Mild cognitive impairment (MCI) is a precursor to dementia marked by declines in communication skills. Whether MCI pathophysiology extends below cerebral cortex to affect speech processing before conscious awareness (brainstem) is unknown. By recording neuroelectric brain activity to speech from brainstem and cortex, we show that MCI hypersensitizes the normal encoding of speech information across the hearing brain. Deficient neural responses to speech (particularly those generated from the brainstem) predicted the presence of MCI with high accuracy and before behavioral deficits. Our findings advance the neurological understanding of MCI by identifying a subcortical biomarker in auditory-sensory processing before conscious awareness, which may be a precursor to declines in speech understanding.
Keywords: auditory evoked potentials; brainstem frequency-following response (FFR); cognitive aging; dementia biomarkers; event-related brain potential (ERPs); speech processing.
Copyright © 2017 the authors 0270-6474/17/373610-11$15.00/0.
Figures
References
-
- Bajo R, Maestú F, Nevado A, Sancho M, Gutierrez R, Campo P, Castellanos NP, Gil P, Moratti S, Pereda E, Del-Pozo F (2010) Functional connectivity in mild cognitive impairment during a memory task: implications for the disconnection hypothesis. J Alzheimers Dis 22:183–193. 10.3233/JAD-2010-100177 - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical