

Understanding the Role of GPCR Heteroreceptor Complexes in Modulating the Brain Networks in Health and Disease
- PMID: 28270751
- PMCID: PMC5318393
- DOI: 10.3389/fncel.2017.00037
Understanding the Role of GPCR Heteroreceptor Complexes in Modulating the Brain Networks in Health and Disease
Abstract
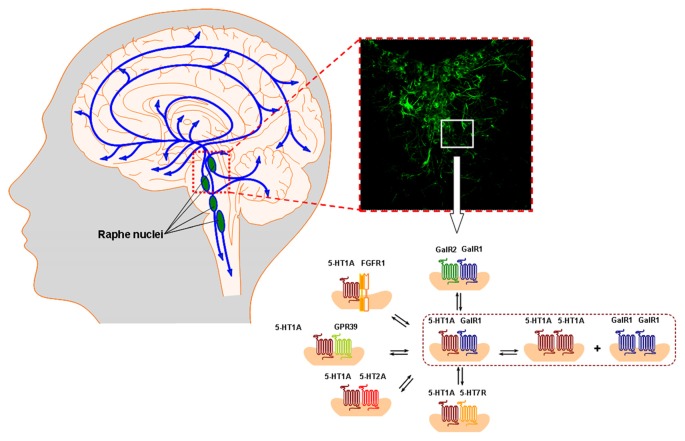
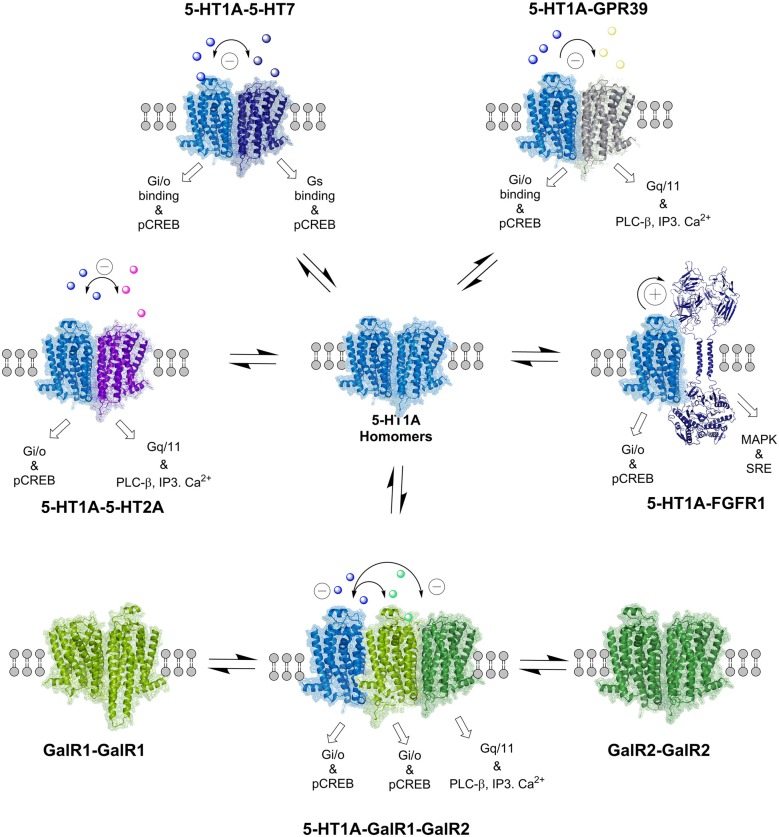
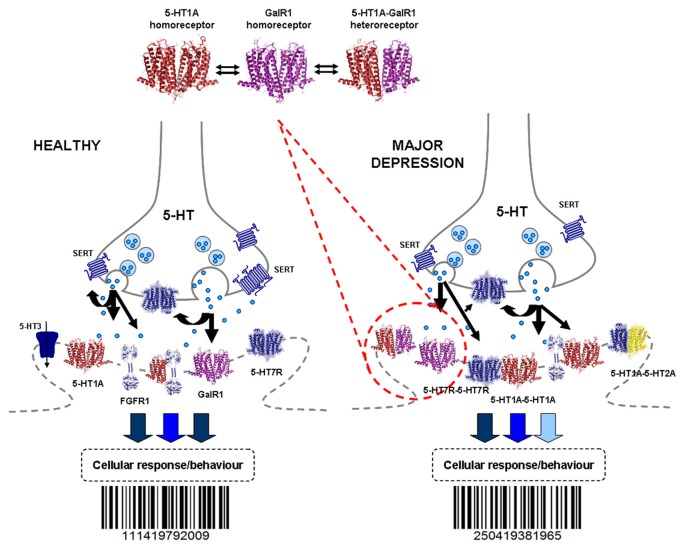
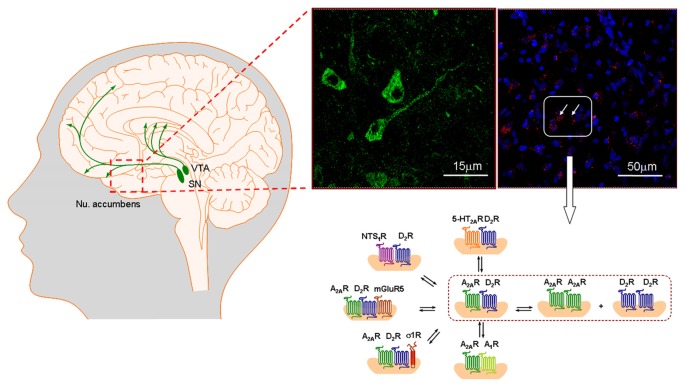
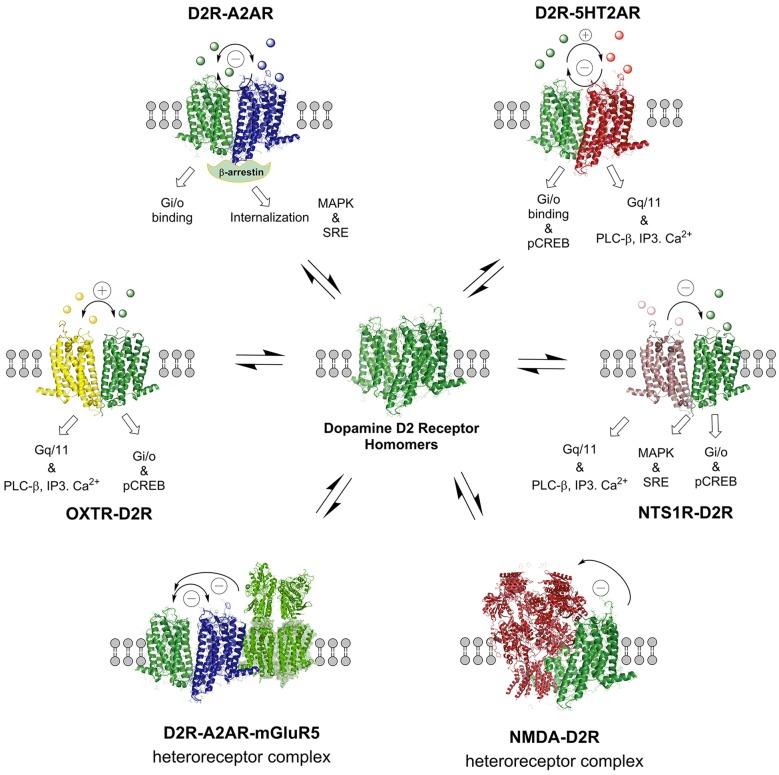
The introduction of allosteric receptor-receptor interactions in G protein-coupled receptor (GPCR) heteroreceptor complexes of the central nervous system (CNS) gave a new dimension to brain integration and neuropsychopharmacology. The molecular basis of learning and memory was proposed to be based on the reorganization of the homo- and heteroreceptor complexes in the postjunctional membrane of synapses. Long-term memory may be created by the transformation of parts of the heteroreceptor complexes into unique transcription factors which can lead to the formation of specific adapter proteins. The observation of the GPCR heterodimer network (GPCR-HetNet) indicated that the allosteric receptor-receptor interactions dramatically increase GPCR diversity and biased recognition and signaling leading to enhanced specificity in signaling. Dysfunction of the GPCR heteroreceptor complexes can lead to brain disease. The findings of serotonin (5-HT) hetero and isoreceptor complexes in the brain over the last decade give new targets for drug development in major depression. Neuromodulation of neuronal networks in depression via 5-HT, galanin peptides and zinc involve a number of GPCR heteroreceptor complexes in the raphe-hippocampal system: GalR1-5-HT1A, GalR1-5-HT1A-GPR39, GalR1-GalR2, and putative GalR1-GalR2-5-HT1A heteroreceptor complexes. The 5-HT1A receptor protomer remains a receptor enhancing antidepressant actions through its participation in hetero- and homoreceptor complexes listed above in balance with each other. In depression, neuromodulation of neuronal networks in the raphe-hippocampal system and the cortical regions via 5-HT and fibroblast growth factor 2 involves either FGFR1-5-HT1A heteroreceptor complexes or the 5-HT isoreceptor complexes such as 5-HT1A-5-HT7 and 5-HT1A-5-HT2A. Neuromodulation of neuronal networks in cocaine use disorder via dopamine (DA) and adenosine signals involve A2AR-D2R and A2AR-D2R-Sigma1R heteroreceptor complexes in the dorsal and ventral striatum. The excitatory modulation by A2AR agonists of the ventral striato-pallidal GABA anti-reward system via targeting the A2AR-D2R and A2AR-D2R-Sigma1R heteroreceptor complex holds high promise as a new way to treat cocaine use disorders. Neuromodulation of neuronal networks in schizophrenia via DA, adenosine, glutamate, 5-HT and neurotensin peptides and oxytocin, involving A2AR-D2R, D2R-NMDAR, A2AR-D2R-mGluR5, D2R-5-HT2A and D2R-oxytocinR heteroreceptor complexes opens up a new world of D2R protomer targets in the listed heterocomplexes for treatment of positive, negative and cognitive symptoms of schizophrenia.
Keywords: G protein-coupled receptor; addiction; depression; dopamine receptor; heteroreceptor complexes; oligomerization; schizophrenia; serotonin receptor.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous