

Antisense Oligonucleotides Used to Target the DUX4 mRNA as Therapeutic Approaches in FaciosScapuloHumeral Muscular Dystrophy (FSHD)
- PMID: 28273791
- PMCID: PMC5368697
- DOI: 10.3390/genes8030093
Antisense Oligonucleotides Used to Target the DUX4 mRNA as Therapeutic Approaches in FaciosScapuloHumeral Muscular Dystrophy (FSHD)
Abstract
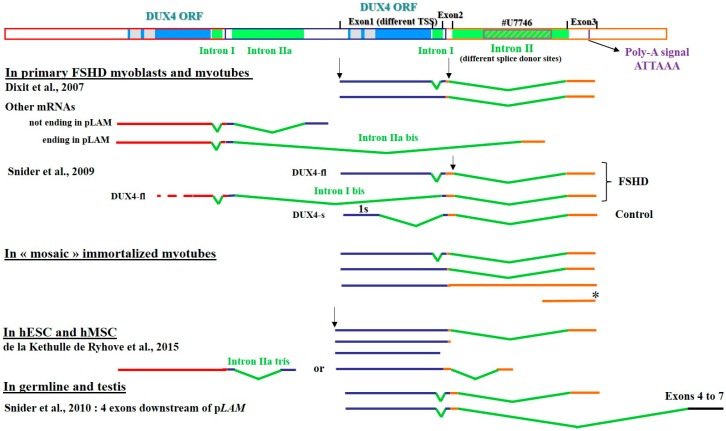
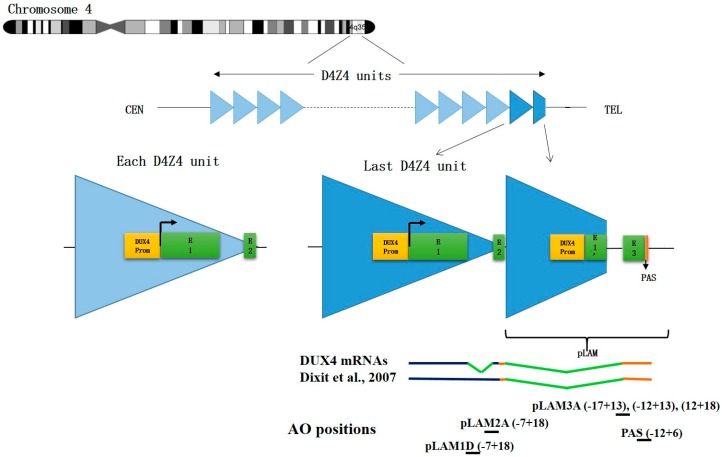
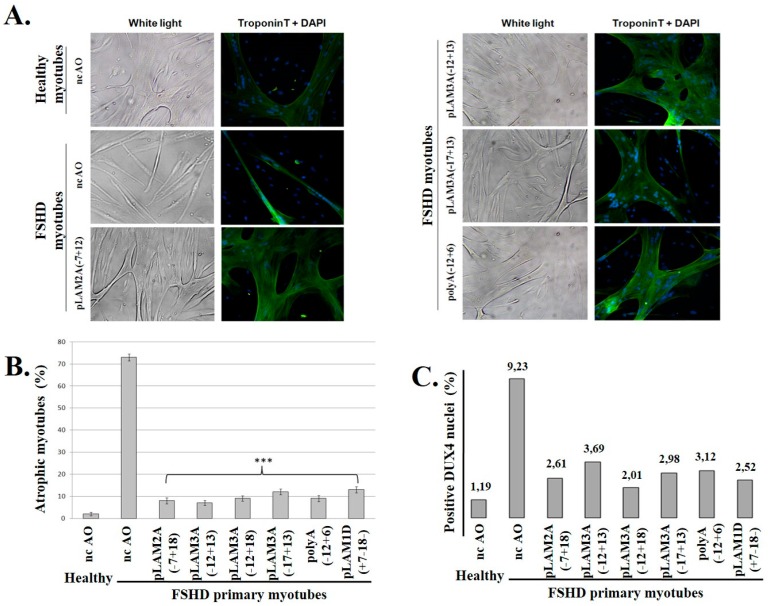
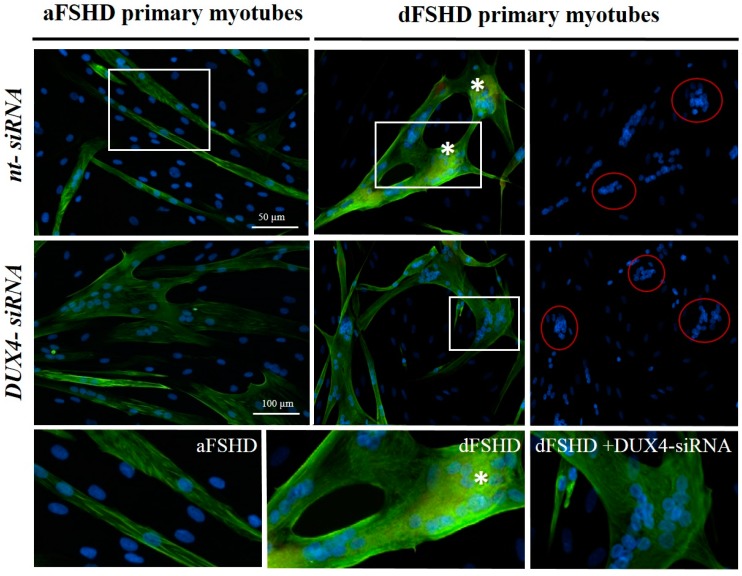
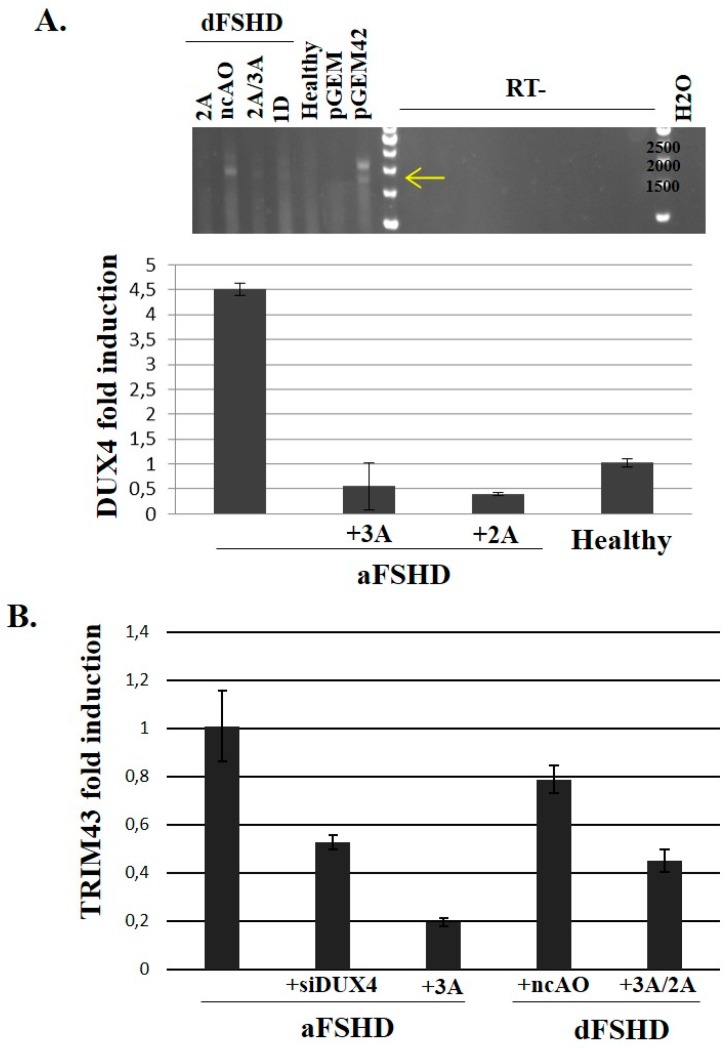
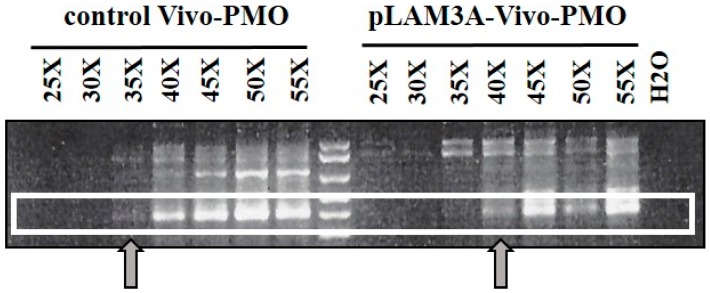
FacioScapuloHumeral muscular Dystrophy (FSHD) is one of the most prevalent hereditary myopathies and is generally characterized by progressive muscle atrophy affecting the face, scapular fixators; upper arms and distal lower legs. The FSHD locus maps to a macrosatellite D4Z4 repeat array on chromosome 4q35. Each D4Z4 unit contains a DUX4 gene; the most distal of which is flanked by a polyadenylation site on FSHD-permissive alleles, which allows for production of stable DUX4 mRNAs. In addition, an open chromatin structure is required for DUX4 gene transcription. FSHD thus results from a gain of function of the toxic DUX4 protein that normally is only expressed in germ line and stem cells. Therapeutic strategies are emerging that aim to decrease DUX4 expression or toxicity in FSHD muscle cells. We review here the heterogeneity of DUX4 mRNAs observed in muscle and stem cells; and the use of antisense oligonucleotides (AOs) targeting the DUX4 mRNA to interfere either with transcript cleavage/polyadenylation or intron splicing. We show in primary cultures that DUX4-targeted AOs suppress the atrophic FSHD myotube phenotype; but do not improve the disorganized FSHD myotube phenotype which could be caused by DUX4c over-expression. Thus; DUX4c might constitute another therapeutic target in FSHD.
Keywords: double homeobox; myopathy; polyadenylation; primary myoblasts; splicing interference.
Conflict of interest statement
E.A., C.V., F.C. and A.B. are inventors in a patent application for antisense agents useful in treating FSHD and targeting either DUX4 or DUX4c (EP patent # 242 62 03, published in the European Patent Bulletin (7 March 2012) and US patent # US20120225034 A, published on 12 September 2012. All authors state that they adhere to Genes′ Publication Ethics Statement.
Figures
References
-
- Orpha.net The portal for rare diseases and orphan drugs. [(accessed on 24 February 2017)]. Available online: http://www.orpha.net.
-
- Deenen J.C.W., Arnts H., van der Maarel S.M., Padberg G.W., Verschuuren J.G.M., Bakker E., Weinreich S.S., Verbeek A.L.M., van Engelen B.G.M. Population-based incidence and prevalence of facioscapulohumeral dystrophy. Neurology. 2014;83:1056–1059. doi: 10.1212/WNL.0000000000000797. - DOI - PMC - PubMed
-
- Turki A., Hayot M., Carnac G., Pillard F., Passerieux E., Bommart S., Raynaud de Mauverger E., Hugon G., Pincemail J., Pietri S., et al. Functional muscle impairment in facioscapulohumeral muscular dystrophy is correlated with oxidative stress and mitochondrial dysfunction. Free Radic. Biol. Med. 2012;53:1068–1079. doi: 10.1016/j.freeradbiomed.2012.06.041. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
