

YidC and SecYEG form a heterotetrameric protein translocation channel
- PMID: 28273911
- PMCID: PMC5427846
- DOI: 10.1038/s41598-017-00109-8
YidC and SecYEG form a heterotetrameric protein translocation channel
Abstract
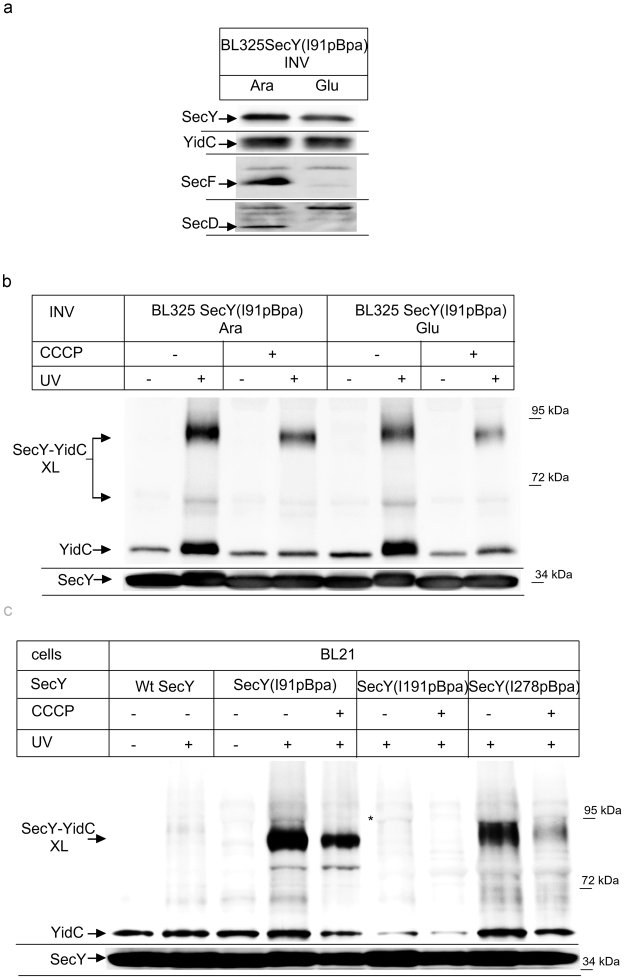
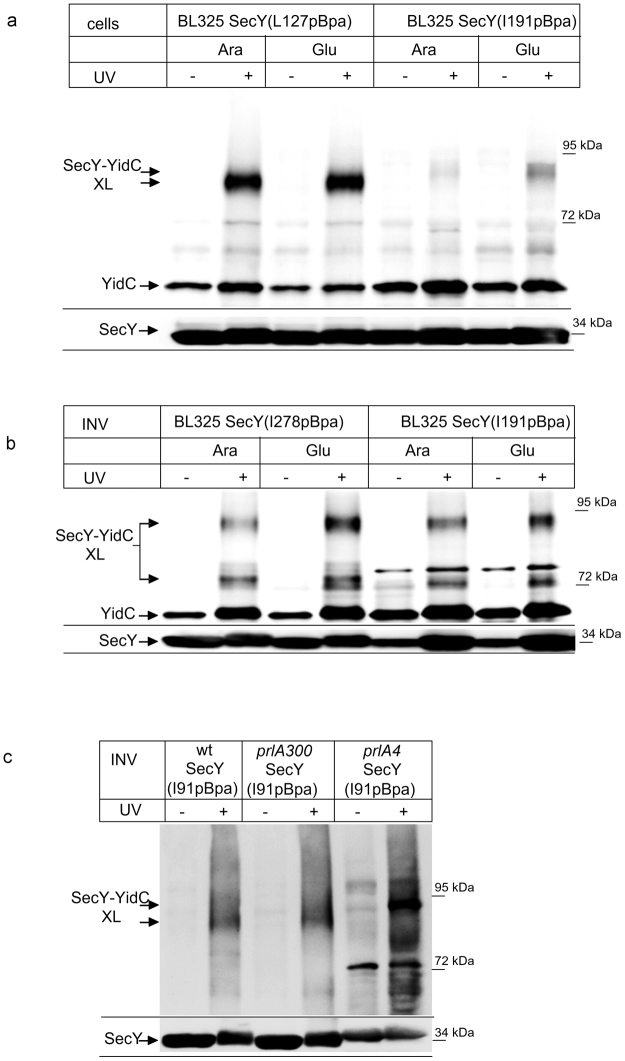
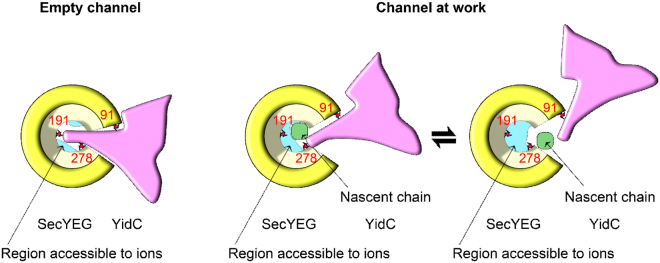
The heterotrimeric SecYEG complex cooperates with YidC to facilitate membrane protein insertion by an unknown mechanism. Here we show that YidC contacts the interior of the SecY channel resulting in a ligand-activated and voltage-dependent complex with distinct ion channel characteristics. The SecYEG pore diameter decreases from 8 Å to only 5 Å for the YidC-SecYEG pore, indicating a reduction in channel cross-section by YidC intercalation. In the presence of a substrate, YidC relocates to the rim of the pore as indicated by increased pore diameter and loss of YidC crosslinks to the channel interior. Changing the surface charge of the pore by incorporating YidC into the channel wall increases the anion selectivity, and the accompanying change in wall hydrophobicity is liable to alter the partition of helices from the pore into the membrane. This could explain how the exit of transmembrane domains from the SecY channel is facilitated by YidC.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
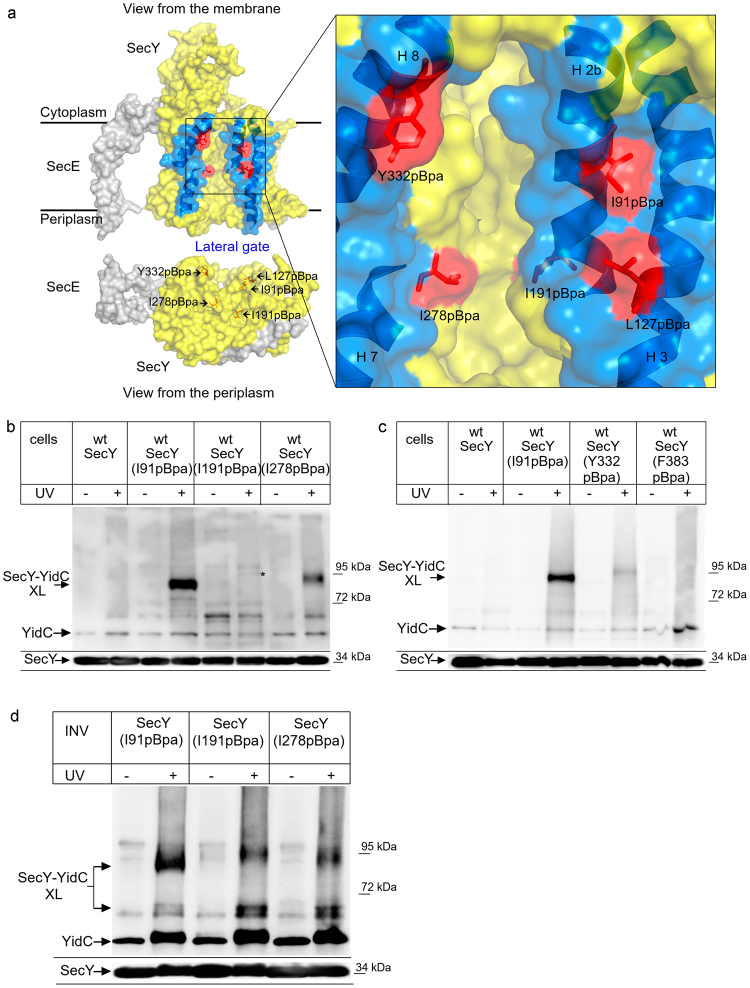
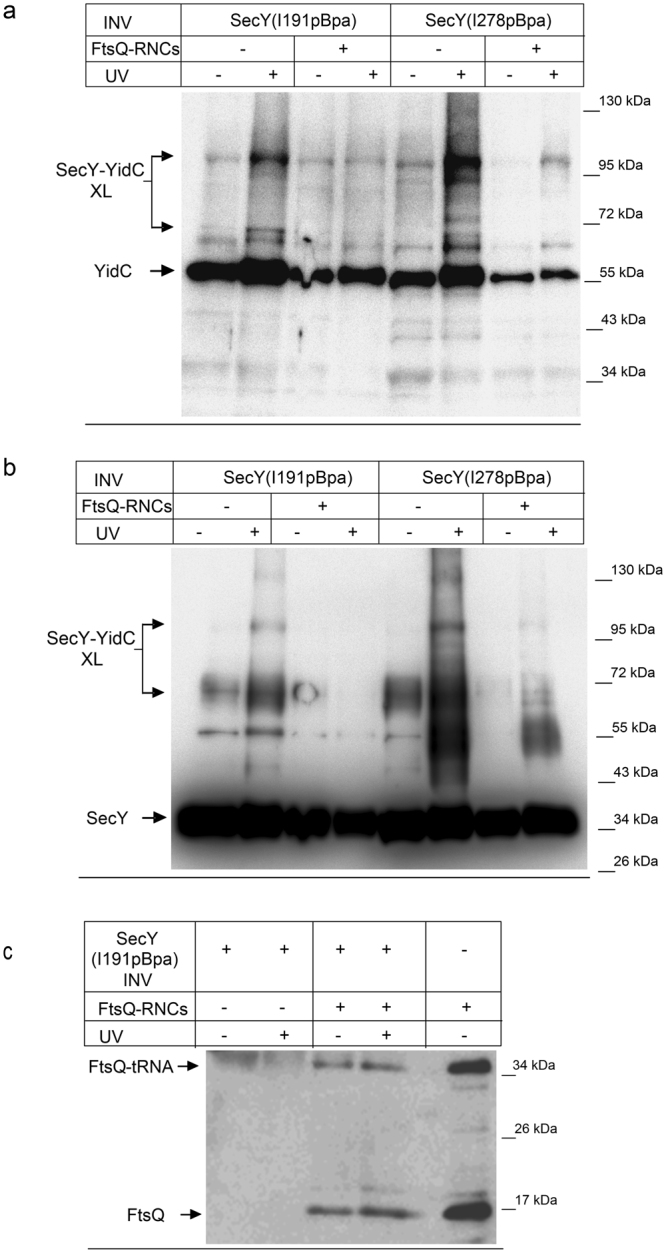
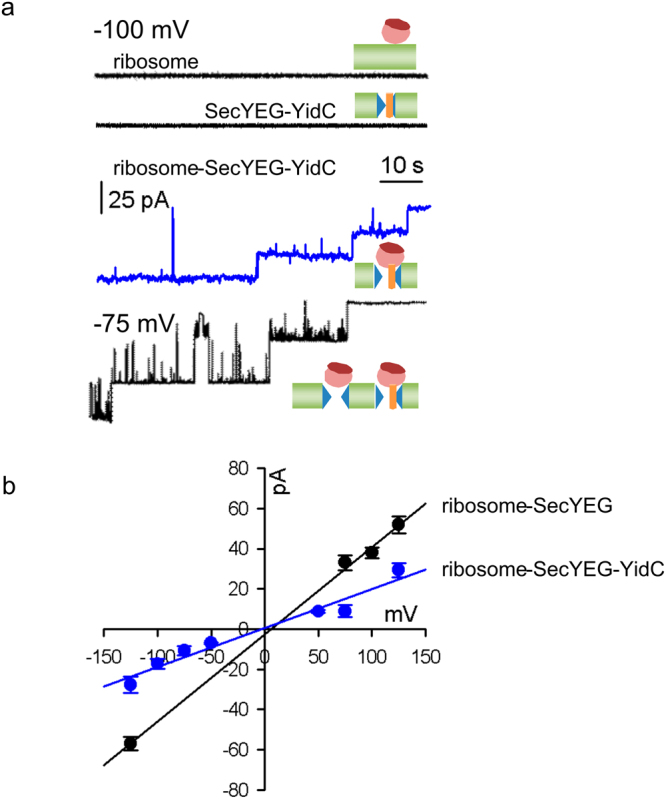
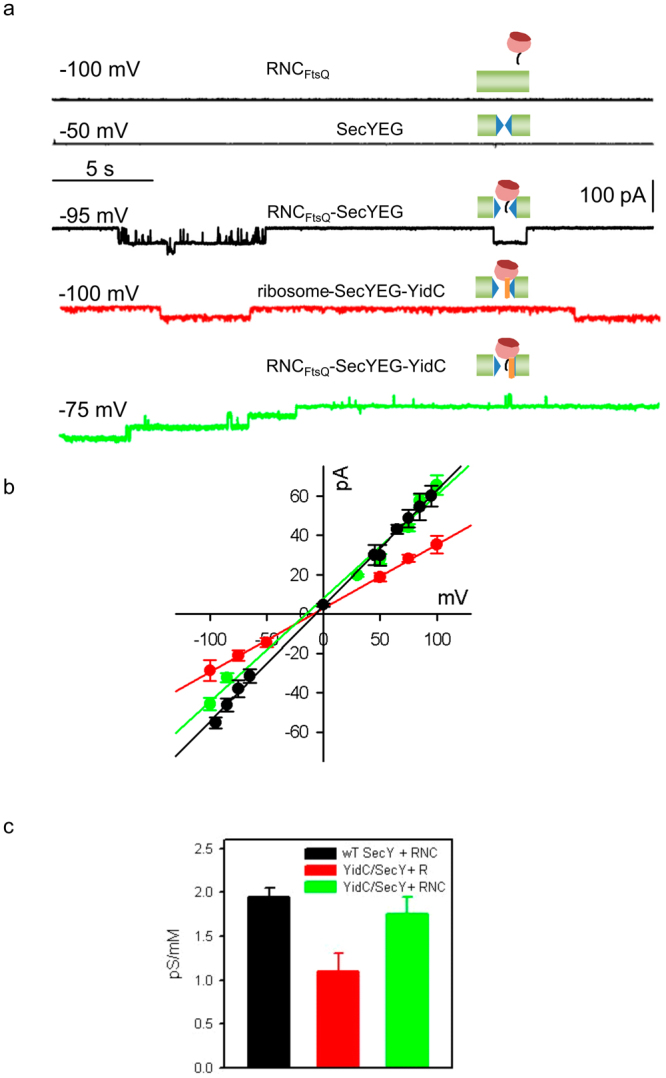
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
