

Pancreatic α- and β-cellular clocks have distinct molecular properties and impact on islet hormone secretion and gene expression
- PMID: 28275001
- PMCID: PMC5358758
- DOI: 10.1101/gad.290379.116
Pancreatic α- and β-cellular clocks have distinct molecular properties and impact on islet hormone secretion and gene expression
Abstract
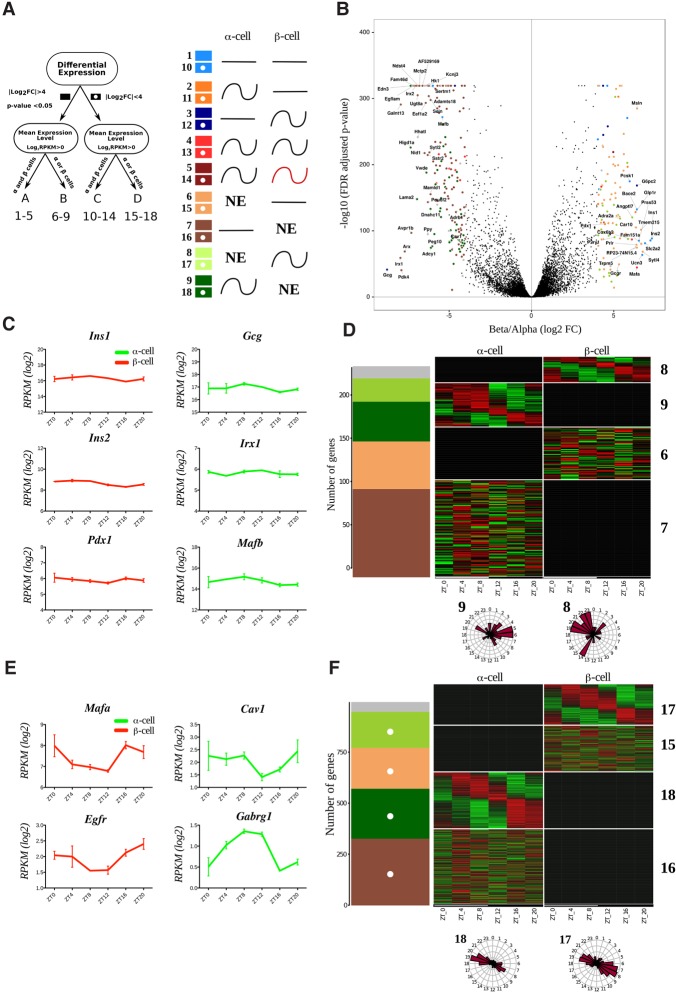
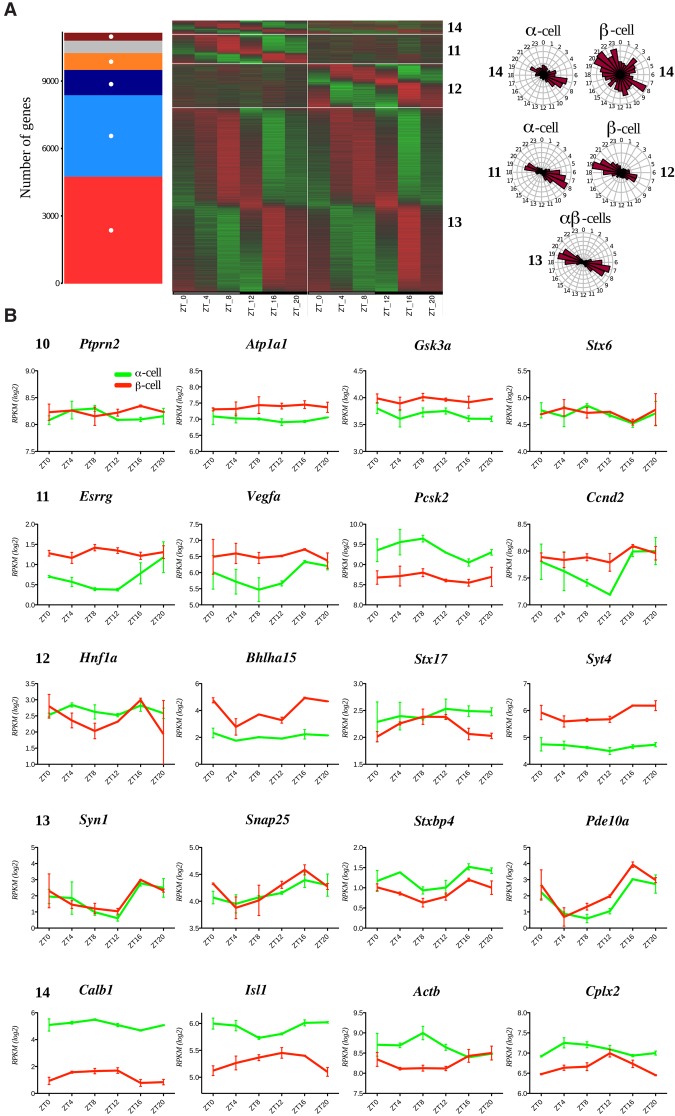
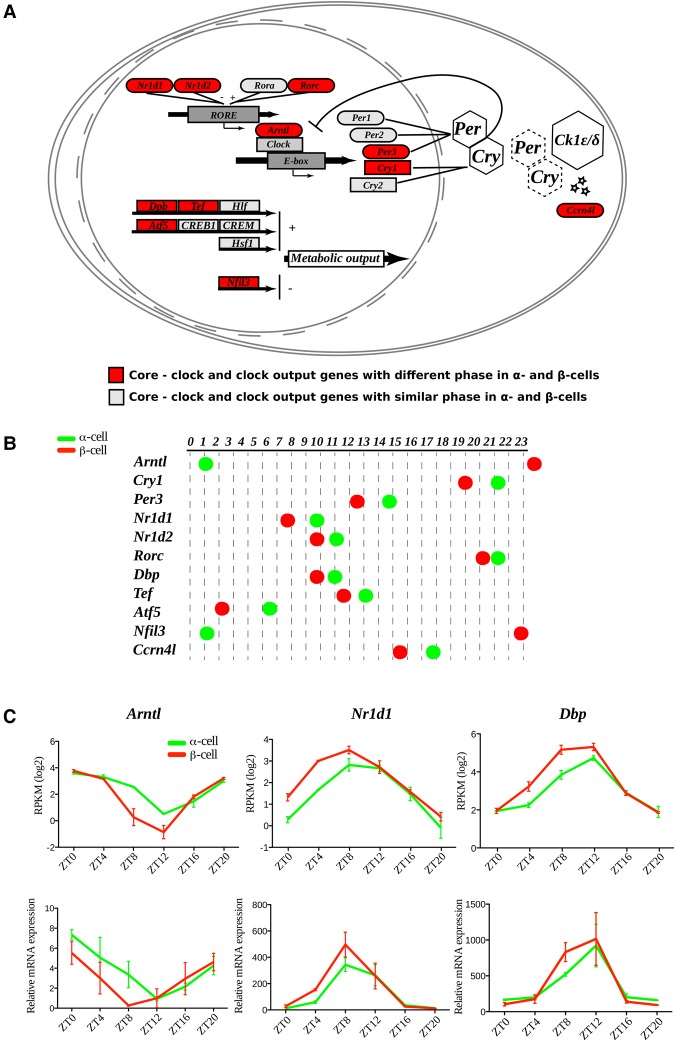
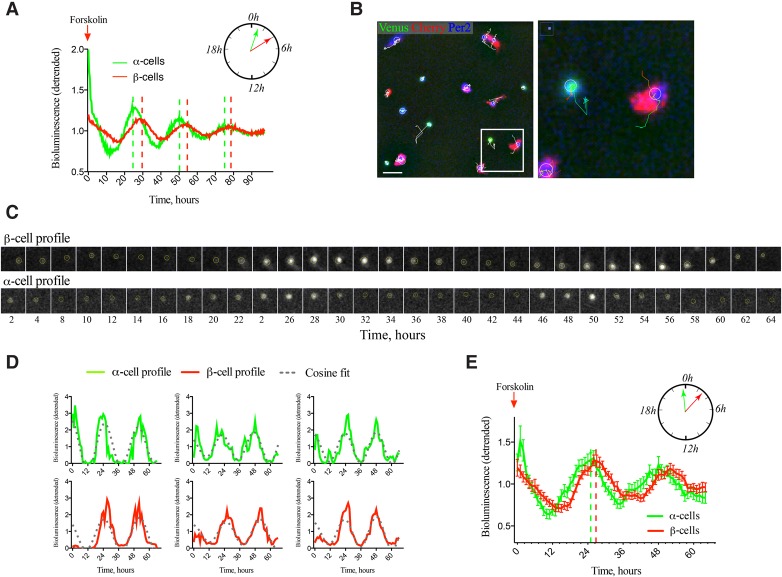
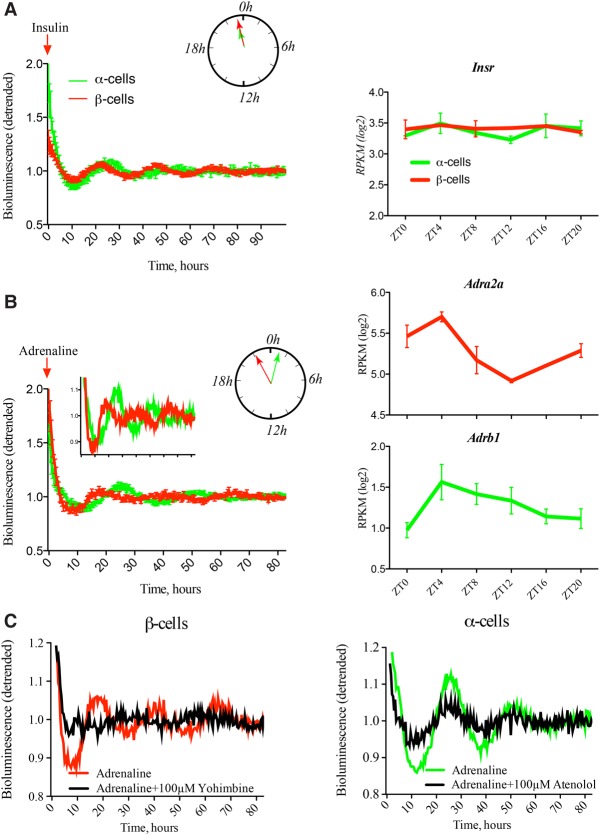
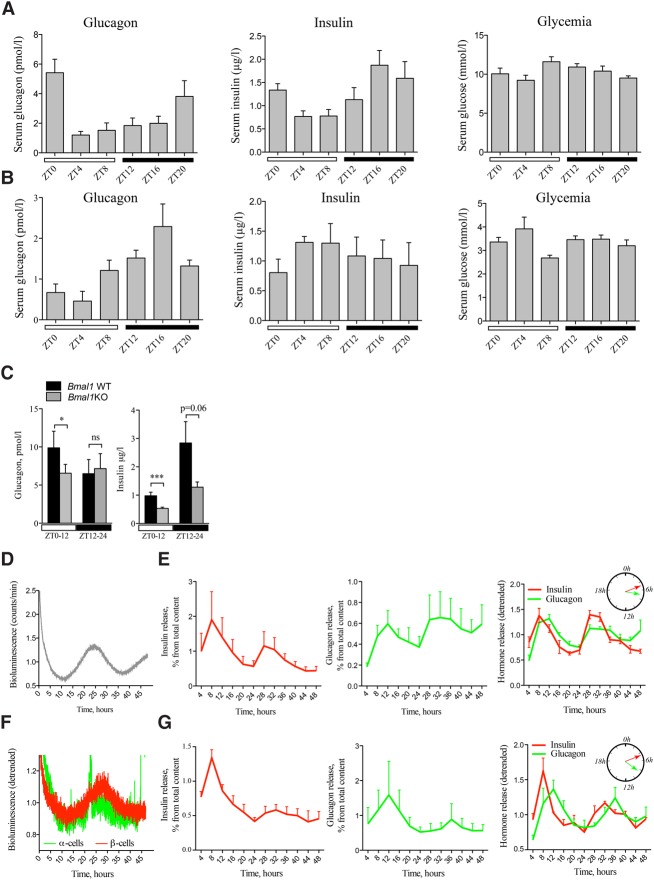
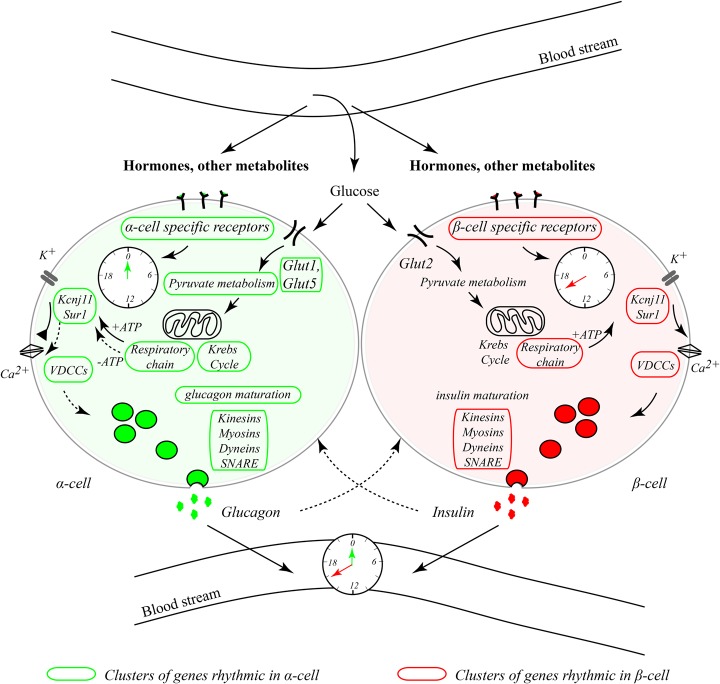
A critical role of circadian oscillators in orchestrating insulin secretion and islet gene transcription has been demonstrated recently. However, these studies focused on whole islets and did not explore the interplay between α-cell and β-cell clocks. We performed a parallel analysis of the molecular properties of α-cell and β-cell oscillators using a mouse model expressing three reporter genes: one labeling α cells, one specific for β cells, and a third monitoring circadian gene expression. Thus, phase entrainment properties, gene expression, and functional outputs of the α-cell and β-cell clockworks could be assessed in vivo and in vitro at the population and single-cell level. These experiments showed that α-cellular and β-cellular clocks are oscillating with distinct phases in vivo and in vitro. Diurnal transcriptome analysis in separated α and β cells revealed that a high number of genes with key roles in islet physiology, including regulators of glucose sensing and hormone secretion, are differentially expressed in these cell types. Moreover, temporal insulin and glucagon secretion exhibited distinct oscillatory profiles both in vivo and in vitro. Altogether, our data indicate that differential entrainment characteristics of circadian α-cell and β-cell clocks are an important feature in the temporal coordination of endocrine function and gene expression.
Keywords: RNA sequencing; circadian clock; insulin/glucagon secretion; mouse α cells and β cells; single-cell bioluminescence–fluorescence time-lapse microscopy.
© 2017 Petrenko et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Comment in
-
Clocks stop sugar shock.Sci Transl Med. 2017 Apr 5;9(384):eaan2772. doi: 10.1126/scitranslmed.aan2772. Sci Transl Med. 2017. PMID: 28381534
References
-
- Asher G, Sassone-Corsi P. 2015. Time for food: the intimate interplay between nutrition, metabolism, and the circadian clock. Cell 161: 84–92. - PubMed
-
- Asher G, Schibler U. 2011. Crosstalk between components of circadian and metabolic cycles in mammals. Cell Metab 13: 125–137. - PubMed
-
- Balsalobre A, Marcacci L, Schibler U. 2000. Multiple signaling pathways elicit circadian gene expression in cultured Rat-1 fibroblasts. Curr Biol 10: 1291–1294. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases