

Meprin Metalloproteases Generate Biologically Active Soluble Interleukin-6 Receptor to Induce Trans-Signaling
- PMID: 28276471
- PMCID: PMC5343444
- DOI: 10.1038/srep44053
Meprin Metalloproteases Generate Biologically Active Soluble Interleukin-6 Receptor to Induce Trans-Signaling
Abstract
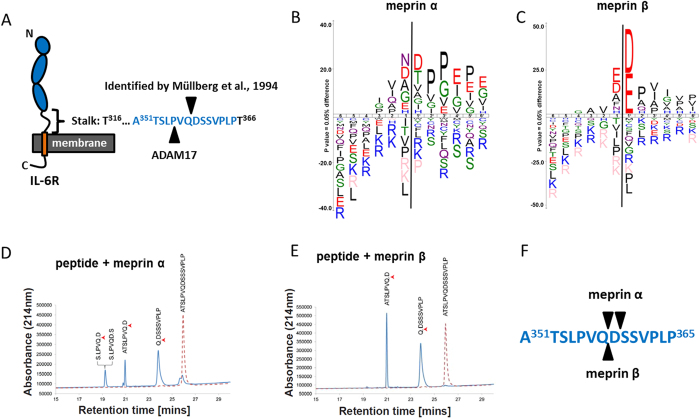
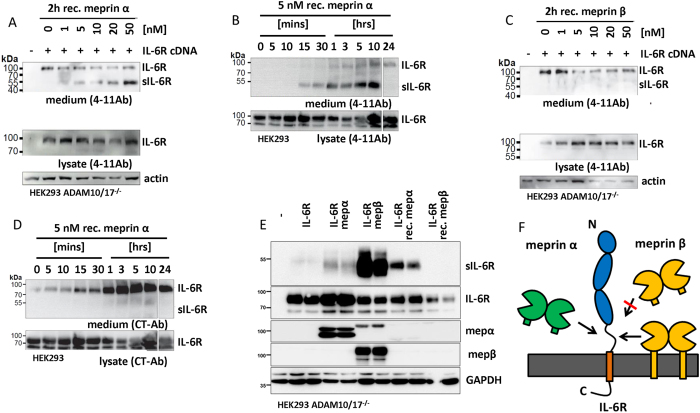
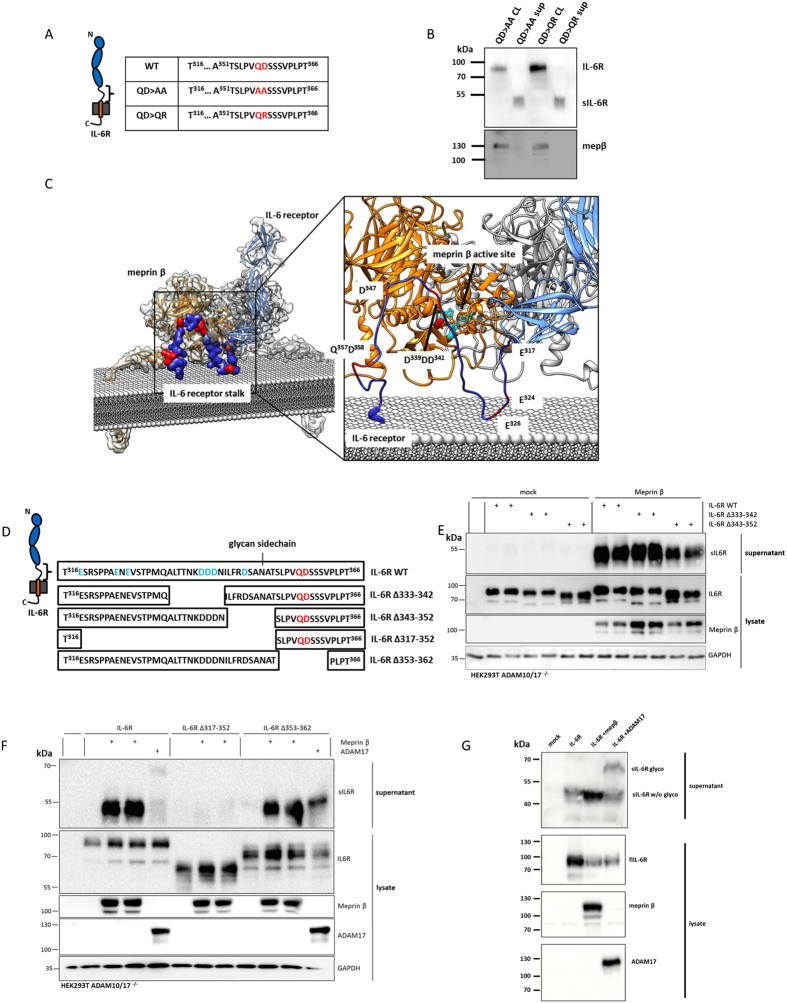
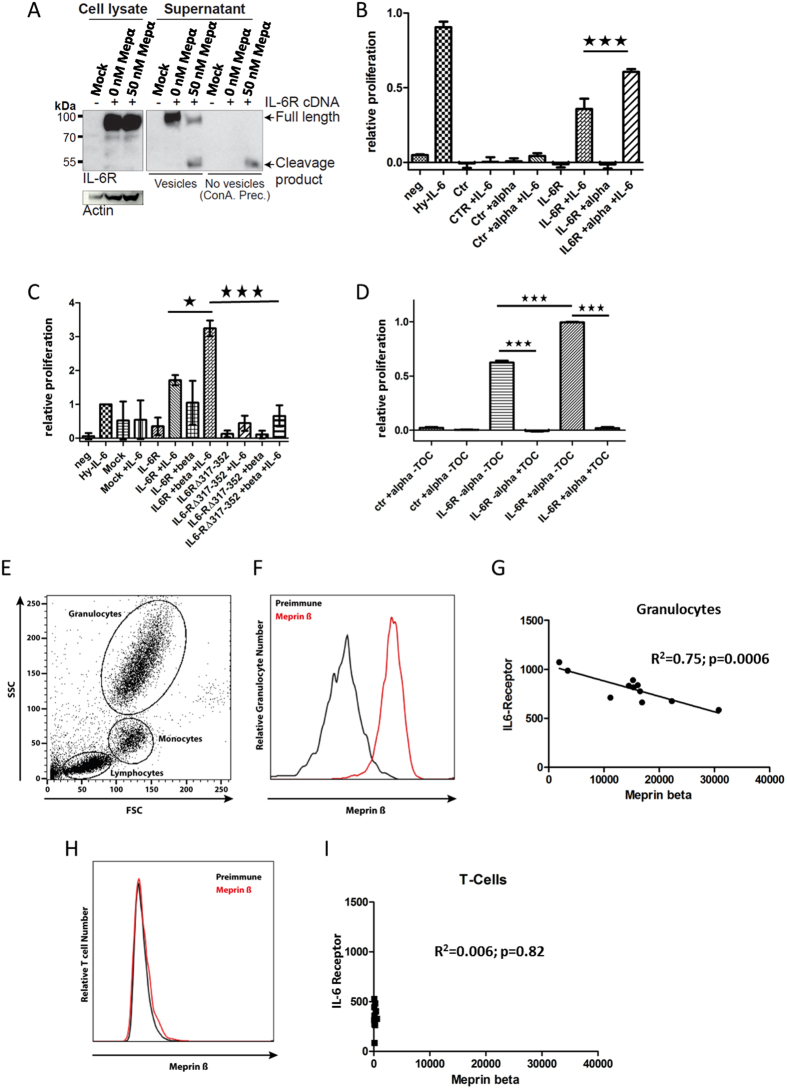
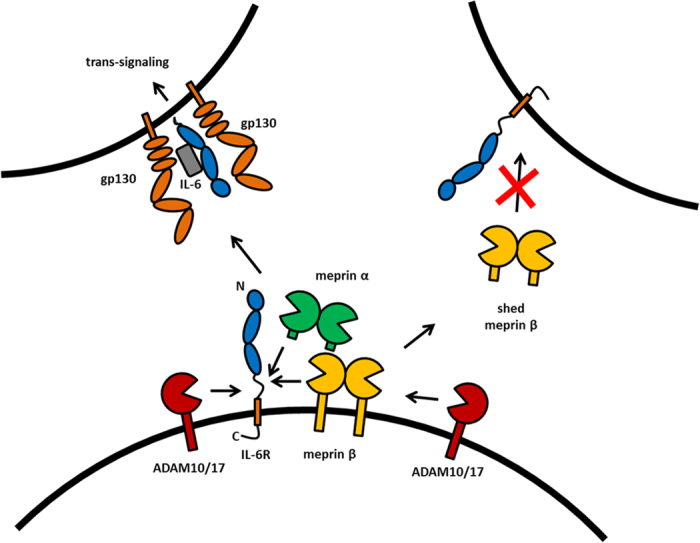
Soluble Interleukin-6 receptor (sIL-6R) mediated trans-signaling is an important pro-inflammatory stimulus associated with pathological conditions, such as arthritis, neurodegeneration and inflammatory bowel disease. The sIL-6R is generated proteolytically from its membrane bound form and A Disintegrin And Metalloprotease (ADAM) 10 and 17 were shown to perform ectodomain shedding of the receptor in vitro and in vivo. However, under certain conditions not all sIL-6R could be assigned to ADAM10/17 activity. Here, we demonstrate that the IL-6R is a shedding substrate of soluble meprin α and membrane bound meprin β, resulting in bioactive sIL-6R that is capable of inducing IL-6 trans-signaling. We determined cleavage within the N-terminal part of the IL-6R stalk region, distinct from the cleavage site reported for ADAM10/17. Interestingly, meprin β can be shed from the cell surface by ADAM10/17 and the observation that soluble meprin β is not capable of shedding the IL-6R suggests a regulatory mechanism towards trans-signaling. Additionally, we observed a significant negative correlation of meprin β expression and IL-6R levels on human granulocytes, providing evidence for in vivo function of this proteolytic interaction.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
